

An arginine switch drives the stepwise activation of β-arrestin by CXCR7
- PMID: 40773476
- PMCID: PMC12331092
- DOI: 10.1371/journal.pbio.3003312
An arginine switch drives the stepwise activation of β-arrestin by CXCR7
Abstract
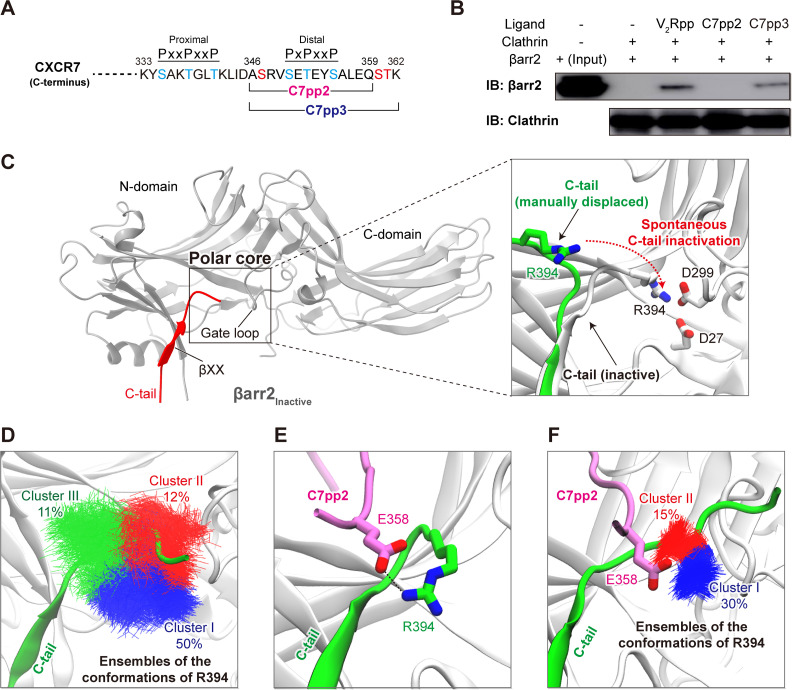
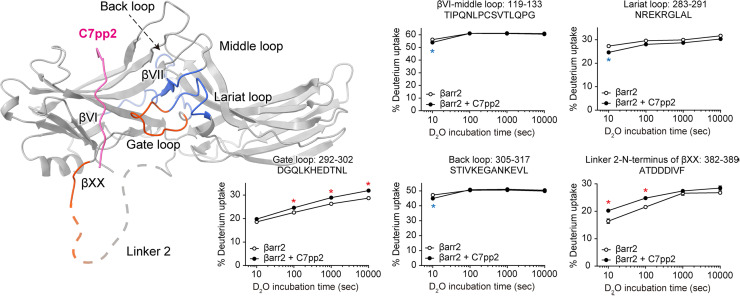
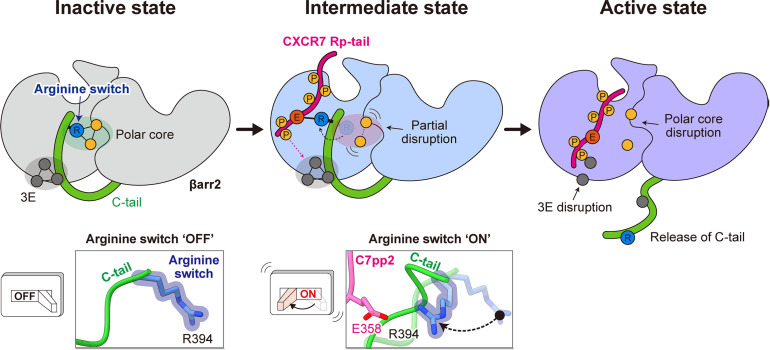
β-arrestins (βarrs) play a crucial role in regulating G protein-coupled receptor (GPCR) signaling and trafficking. Canonically, interactions of βarr with the phosphorylated intracellular GPCR-tail induce a multi-step conformational transition that results in the activation of βarr. Depending on the specific interaction pattern with the receptor, βarrs adopt multiple conformational states, each tightly linked to a specific functional outcome of βarr recruitment. Despite its physiological relevance, the structural determinants of βarr activation remain poorly understood. Using a combination of molecular dynamics simulations, biochemical and cell-based experiments, we reveal how specific interactions with a chemokine receptor 7 (CXCR7) promote the unbinding of the βarr2 C-tail-a crucial step in arrestin activation. Importantly, we observe that the expulsion of the C-tail is promoted by the displacement of a conserved arginine residue (Arg394) within the βarr polar core, which we dub "the arginine switch." Our study uncovers a role for the arginine switch that, upon engagement, destabilizes the polar core as a crucial step in the CXCR7-induced βarr activation.
Copyright: © 2025 Ji et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
