

This is a preprint.
Human iPSC derived alveolar macrophages reveal macrophage subtype specific functions of itaconate in M. tuberculosis host defense
- PMID: 40777401
- PMCID: PMC12330731
- DOI: 10.1101/2025.07.23.664455
Human iPSC derived alveolar macrophages reveal macrophage subtype specific functions of itaconate in M. tuberculosis host defense
Abstract
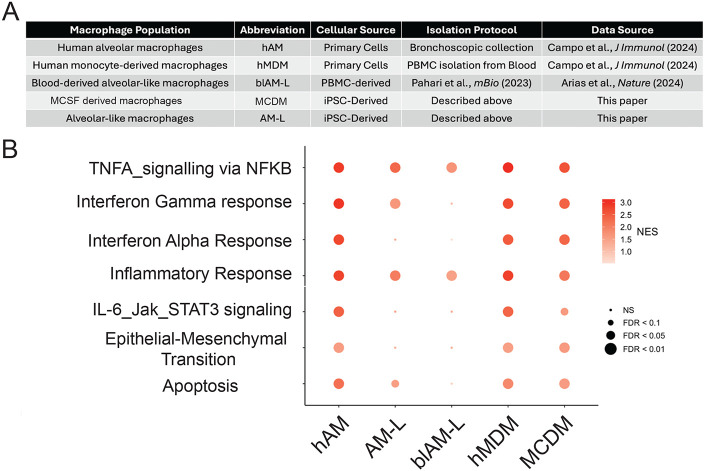
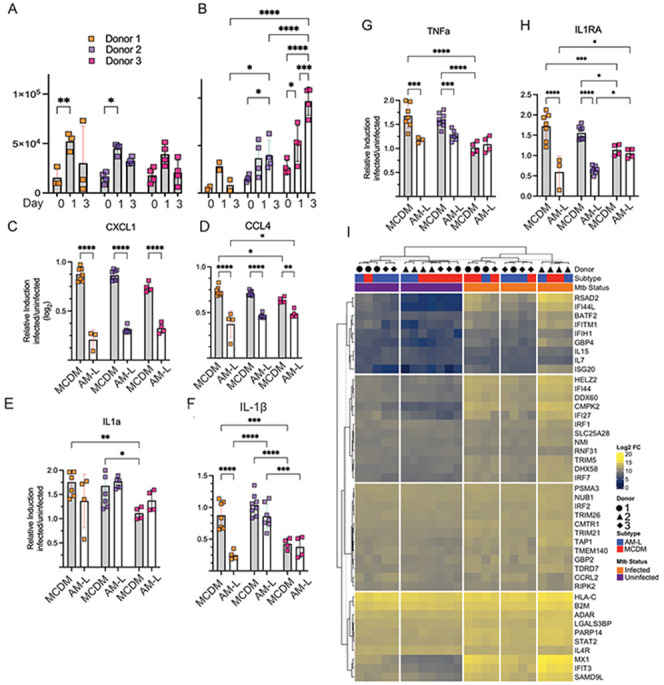
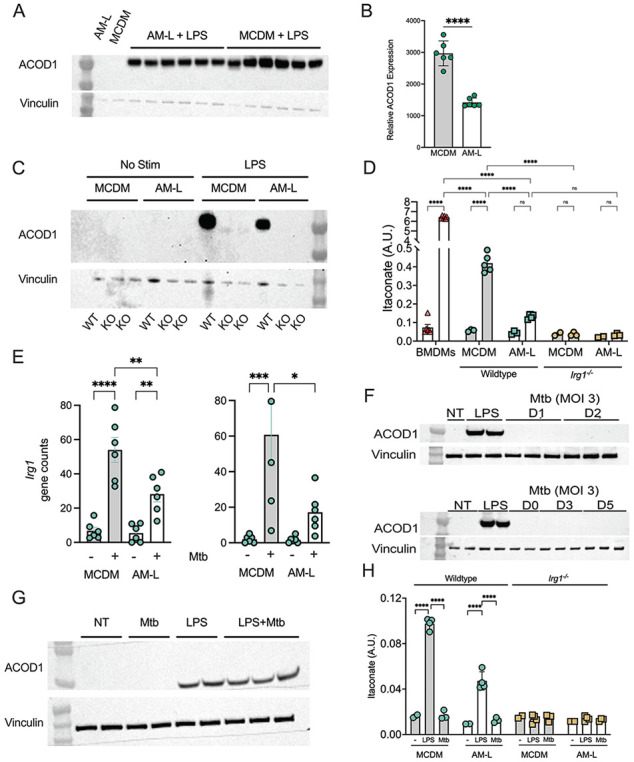
Mycobacterium tuberculosis (Mtb) must survive within multiple macrophage populations during infection, including alveolar macrophages (AM) and recruited inflammatory macrophages. In mice, itaconate, produced in macrophages by ACOD1 mediated decarboxylation of aconitate, has direct antimicrobial activity, modulates inflammatory cytokines, and is required for resistance to M. tuberculosis (Mtb) infection. The role of itaconate in human macrophages is less clear and whether itaconate mediates distinct effects in macrophage subtypes is unknown. Here, we investigated the role of itaconate in human iPSC-derived macrophages, either induced by GM-CSF to resemble alveolar macrophages (AM-Like cells), or treated with M-CSF to generate control macrophages (MCDM cells). Both types of human macrophages produce substantially less itaconate than mouse macrophages and AM-Ls produced 4-fold less itaconate than MCDMs. Surprisingly, ACOD1 deficient AM-L macrophages, but not MCDM macrophages, were permissive for Mtb growth. Moreover, itaconate functioned to dampen the Mtb induced inflammatory response in MCDMs, but not AM-L macrophages, affecting both the Type I IFN and TNF pathways. These results indicate that itaconate is involved in human macrophage responses to TB, with distinct roles in different macrophage subsets. These results also show that genetically tractable hiPSC-derived macrophages are a robust and versatile model to dissect cellular host pathogen interactions.
Conflict of interest statement
Conflict of interest MSG reports consulting fees and equity from Vedanta Biosciences and consulting fees from Fimbrion. KADM reports consulting fees from EuropaDX for bioinformatic tool development not related to this project.
Figures


References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials