

This is a preprint.
Adrenergic signaling coordinates distant and local responses to amputation in axolotl
- PMID: 40777403
- PMCID: PMC12330692
- DOI: 10.1101/2021.12.29.474455
Adrenergic signaling coordinates distant and local responses to amputation in axolotl
Update in
-
Adrenergic signaling coordinates distant and local responses to amputation in axolotl.Cell. 2025 Oct 24:S0092-8674(25)01125-0. doi: 10.1016/j.cell.2025.09.025. Online ahead of print. Cell. 2025. PMID: 41138727
Abstract
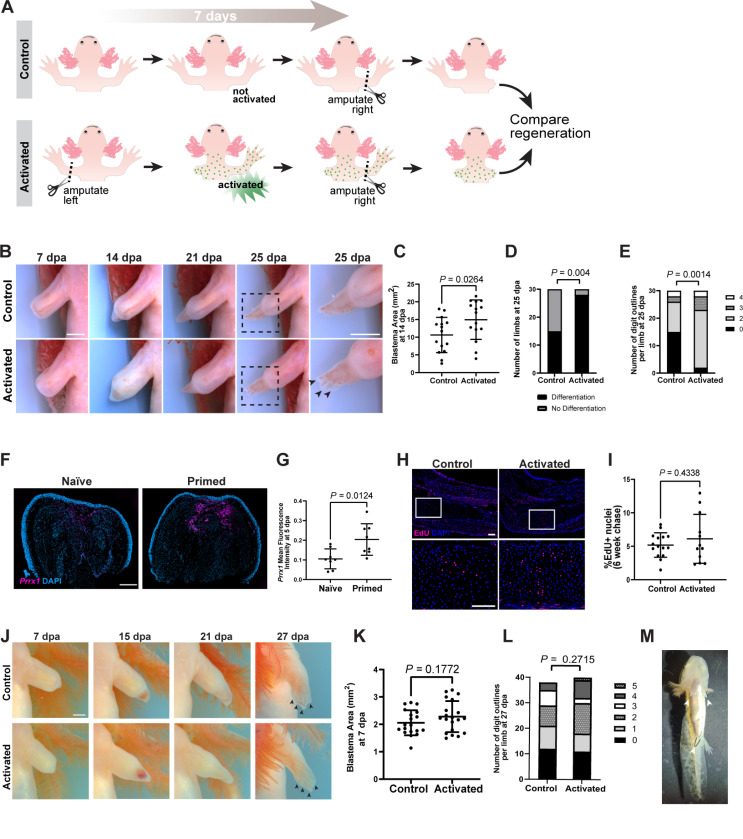
Many species regenerate lost body parts following amputation. Most limb regeneration research has focused on the immediate injury site. Meanwhile, body-wide injury responses remain largely unexplored but may be critical for regeneration. Here, we discovered a role for the sympathetic nervous system in stimulating a body-wide stem cell activation response to amputation that drives enhanced limb regeneration in axolotls. This response is mediated by adrenergic signaling, which coordinates distant cellular activation responses via the α2A-adrenergic receptor, and local regeneration responses via β-adrenergic receptors. Both α2A- and β-adrenergic signaling act upstream of mTOR signaling. Notably, systemically-activated axolotls regenerate limbs faster than naïve animals, suggesting a potential selective advantage in environments where injury from cannibalism or predation is common. This work challenges the predominant view that cellular responses underlying regeneration are confined to the injury site and argues instead for body-wide cellular priming as a foundational step that enables localized tissue regrowth.
Keywords: amputation; limb; mTOR; noradrenaline; norepinephrine; peripheral nervous system; progenitor cells; regeneration; stem cells; systemic responses.
Conflict of interest statement
J.L.W. is a co-founder of Animate Biosciences. I.M.C. consults for GSK Pharmaceuticals, and his lab has received sponsored research from Moderna and AbbVie/Allergan. J.D.B. holds patents related to ATAC-seq, consults for the Treehouse Family Foundation and is a Scientific Advisory Board member of Camp4 and seqWell. M.V.P. is a co-founder of Amplifica Holdings Group, consults for L’Oreal and ODDITY, and his lab has received sponsored research from L’Oreal and AbbVie/Allergan. Other authors declare no competing interests.
Figures






References
-
- Moreno G. (1989). Behavioral and Physiological Differentiation between the Color Morphs of the Salamander, Plethodon cinereus. Journal of Herpetology 23, 335–341. 10.2307/1564043. - DOI
-
- Group, I.S.A.S. (2020). Ambystoma mexicanum. The IUCN Red List of Threatened Species 2020: e.T1095A53947343.
-
- Miller B.T., and Miller J.L. (2005). Prevalence of physical abnormalities in eastern hellbender (Cryptobranchus alleganiensis alleganiensis) populations of Middle Tennessee. Southeastern Naturalist 4, 513–520.
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous