

Brain transcriptomics highlight abundant gene expression and splicing alterations in non-neuronal cells in aFTLD-U
- PMID: 40783910
- PMCID: PMC12336083
- DOI: 10.1007/s00401-025-02919-x
Brain transcriptomics highlight abundant gene expression and splicing alterations in non-neuronal cells in aFTLD-U
Abstract
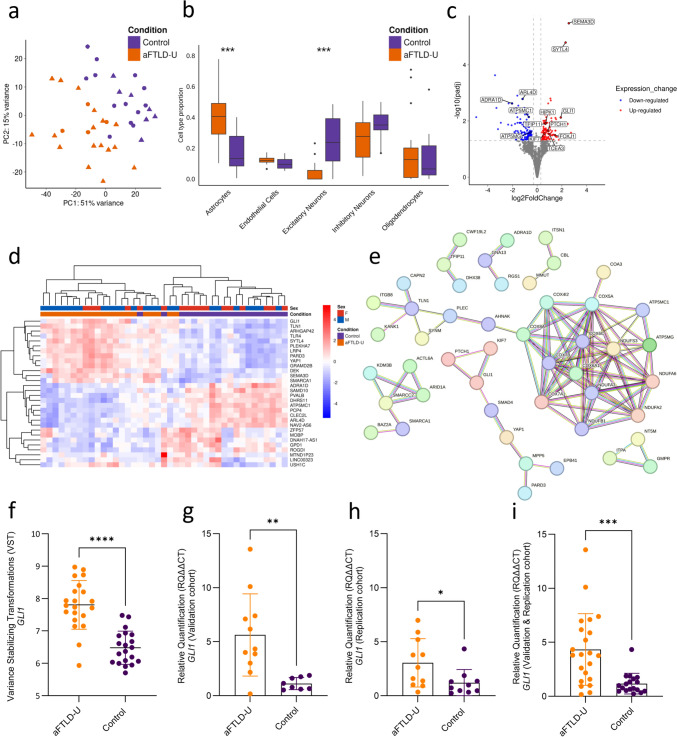
Atypical frontotemporal lobar degeneration with ubiquitin-positive inclusions (aFTLD-U) is a rare cause of frontotemporal lobar degeneration (FTLD), characterized postmortem by neuronal inclusions of the FET family of proteins (FTLD-FET). The recent discovery of TAF15 amyloid filaments in aFTLD-U brains represents a significant step toward improved diagnostic and therapeutic strategies. However, our understanding of the etiology of this FTLD subtype remains limited, which severely hampers translational research efforts. To explore the transcriptomic changes in aFTLD-U, we performed bulk RNA sequencing on the frontal cortex tissue of 21 aFTLD-U patients and 20 control individuals. Cell-type deconvolution revealed loss of excitatory neurons and a higher proportion of astrocytes in aFTLD-U relative to controls. Differential gene expression and co-expression network analysis, adjusted for the shift in cell-type proportions, showed dysregulation of mitochondrial pathways, transcriptional regulators, and upregulation of the Sonic hedgehog (Shh) pathway, including the GLI1 transcription factor, in aFTLD-U. Overall, oligodendrocyte and astrocyte-enriched genes were significantly over-represented among the differentially expressed genes. Differential splicing analysis confirmed the dysregulation of non-neuronal cell types with significant splicing alterations, particularly in oligodendrocyte-enriched genes, including myelin basic protein (MBP), a crucial component of myelin. Immunohistochemistry in frontal cortex brain tissue also showed reduced myelin levels in aFTLD-U patients compared to controls. Together, these findings highlight a central role for glial cells, particularly astrocytes and oligodendrocytes, in the pathogenesis of aFTLD-U, with disruptions in mitochondrial activity, RNA metabolism, Shh signaling, and myelination as possible disease mechanisms. This study offers the first transcriptomic insight into aFTLD-U and presents new avenues for research into FTLD-FET.
Keywords: Glial cells; Mitochondria; Sonic hedgehog signaling; Splicing; Transcriptomics; aFTLD-U.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Conflict of interest: Dr. Rademakers receives invention royalties from a patent related to progranulin. Dr. Mackenzie is a member of the Scientific Advisory Board of Prevail Therapeutics and receives invention royalties from a patent related to progranulin. Drs. Rademakers and De Coster are inventors on a patent filed concerning diagnostic applications in aFTLD-U. Ethical approval and consent to participate: This study was approved by the University of Antwerp and Mayo Clinic Institutional Review Boards. The collection of tissue samples was approved by the Mayo Clinic Institutional Review Board. All autopsies were obtained after consent by the legal next-of-kin or someone legally authorized to make this decision.
Figures


References
MeSH terms
Grants and funding
- 12ASR24N/Fund for Scientific Research Flanders
- R35 NS097261/NS/NINDS NIH HHS/United States
- P30AG062422/National Institutes of Health,United States
- P30 AG062422/AG/NIA NIH HHS/United States
- P30 AG066530/AG/NIA NIH HHS/United States
- R35NS097261/National Institutes of Health,United States
- U19AG063911/National Institutes of Health,United States
- P30 AG066468/AG/NIA NIH HHS/United States
- 74580/CAPMC/ CIHR/Canada
- R21 NS110994/NS/NINDS NIH HHS/United States
- U19 AG063911/AG/NIA NIH HHS/United States
- DOCPRO4/University of Antwerp
- 1176607/National Health and Medical Research Council of Australia Senior Leadership Fellowship
- R21NS110994/National Institutes of Health,United States
- P30AG066468/National Institutes of Health,United States
LinkOut - more resources
Full Text Sources
Miscellaneous
