

Frequency-Dependent Ecological Interactions Increase the Prevalence, and Shape the Distribution, of Preexisting Drug Resistance
- PMID: 40786663
- PMCID: PMC12333505
- DOI: 10.1103/prxlife.2.023010
Frequency-Dependent Ecological Interactions Increase the Prevalence, and Shape the Distribution, of Preexisting Drug Resistance
Abstract
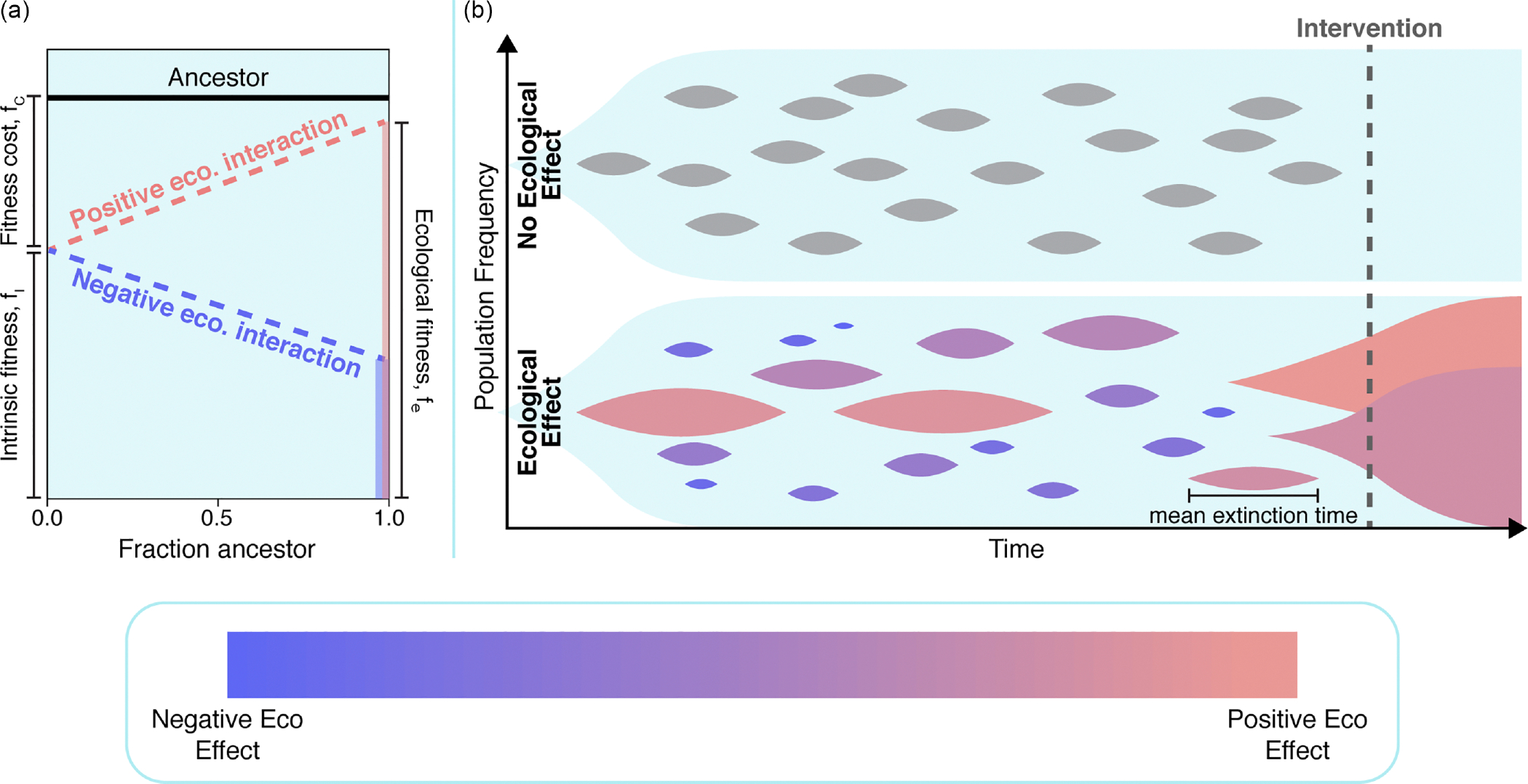
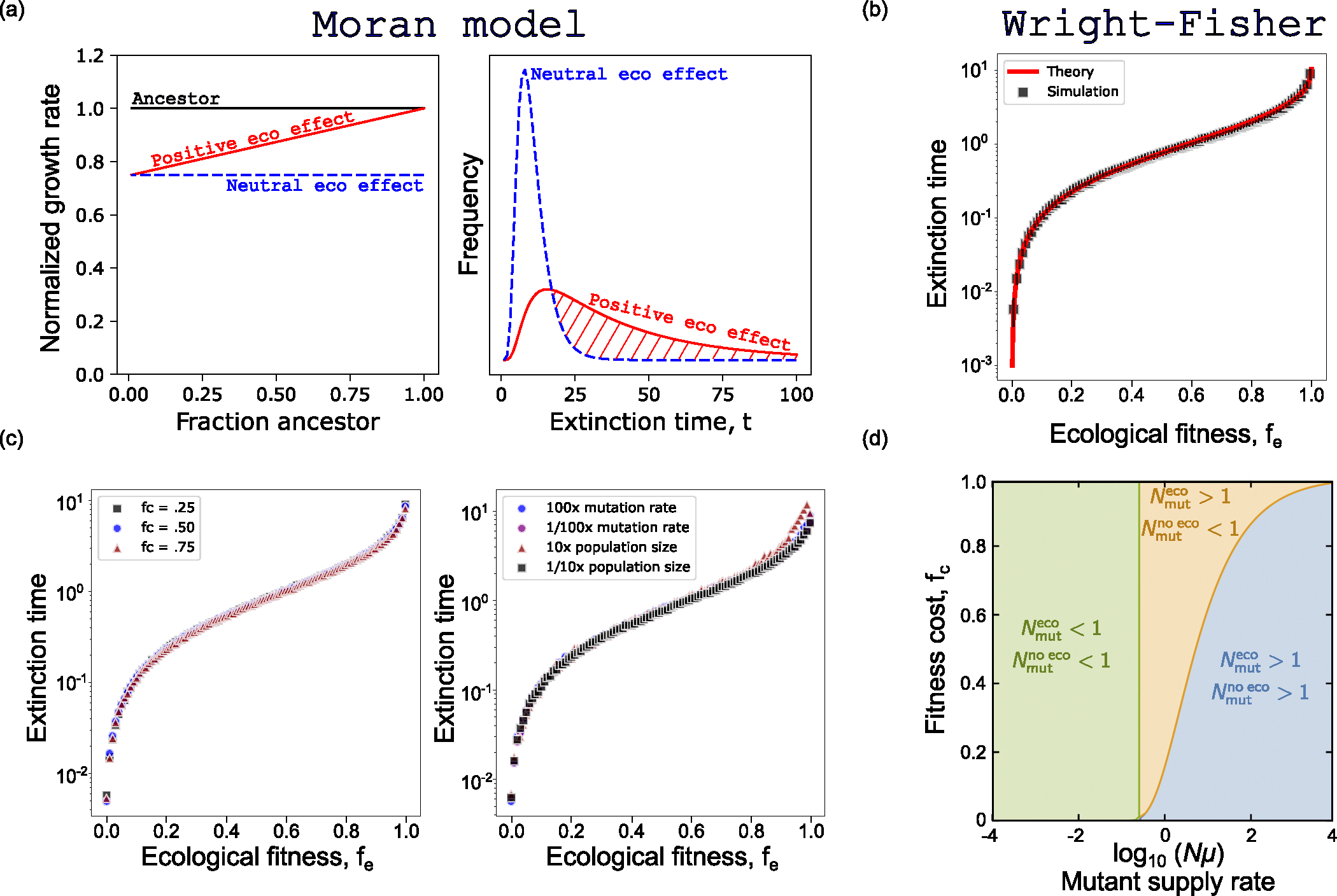
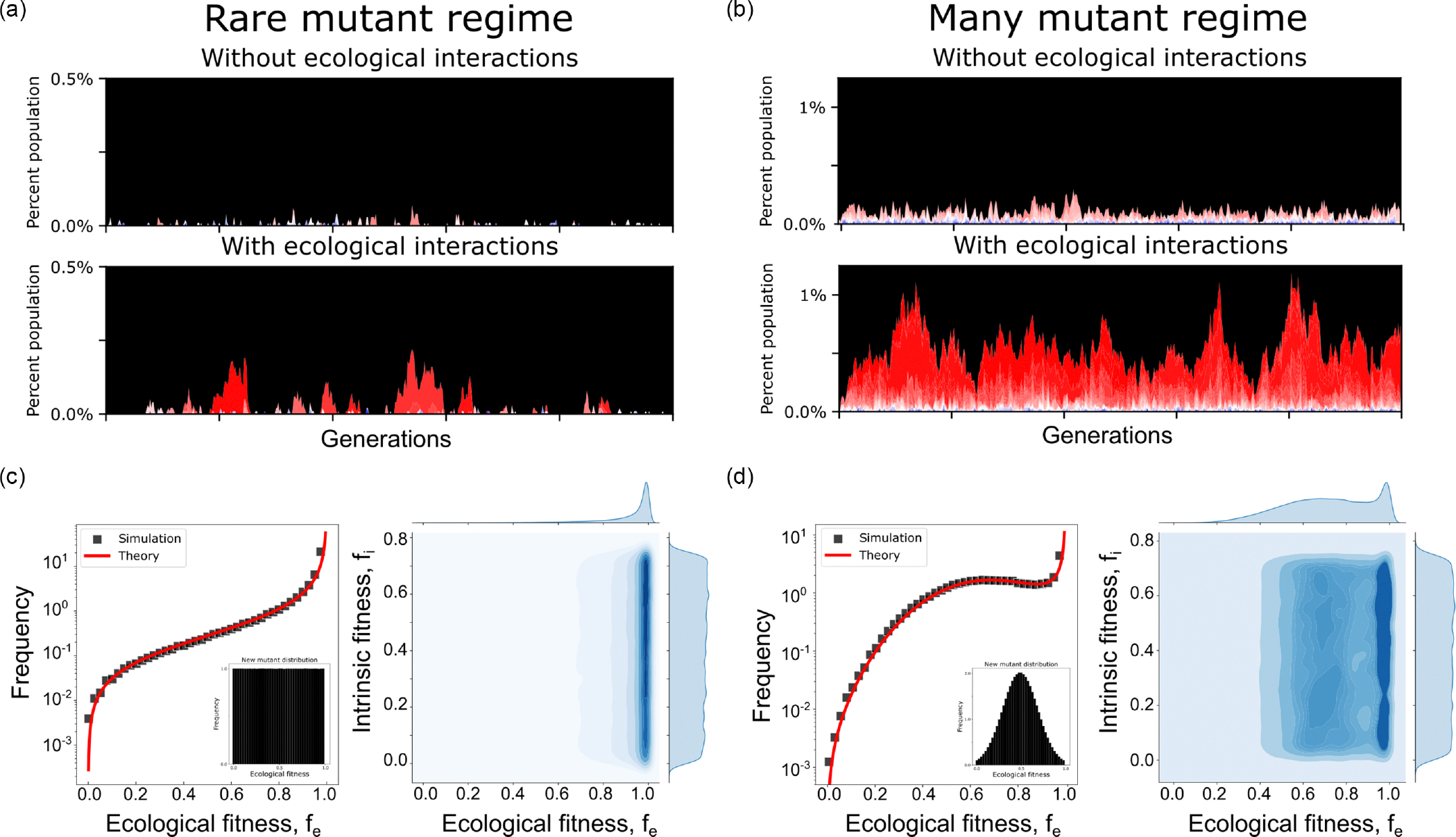
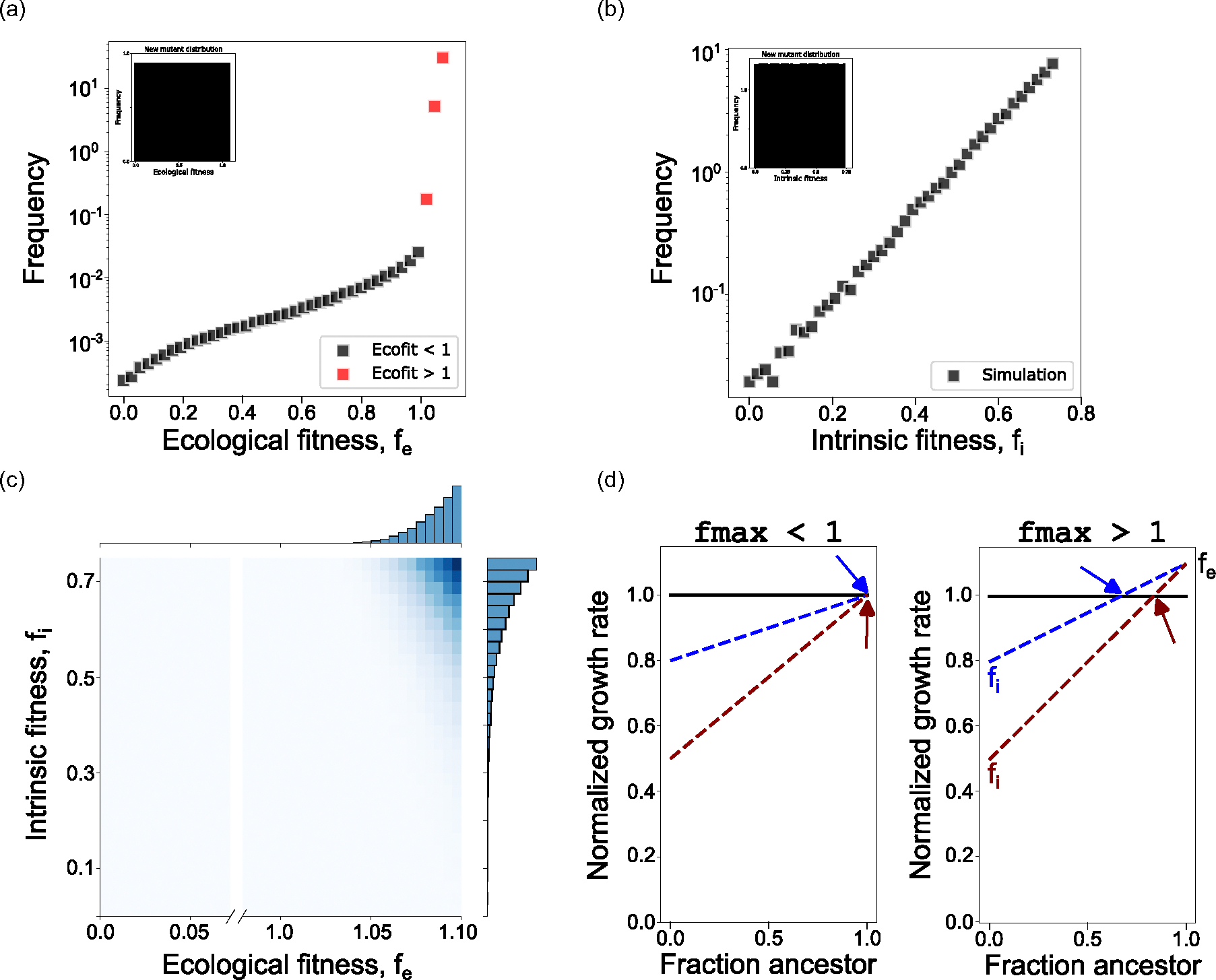
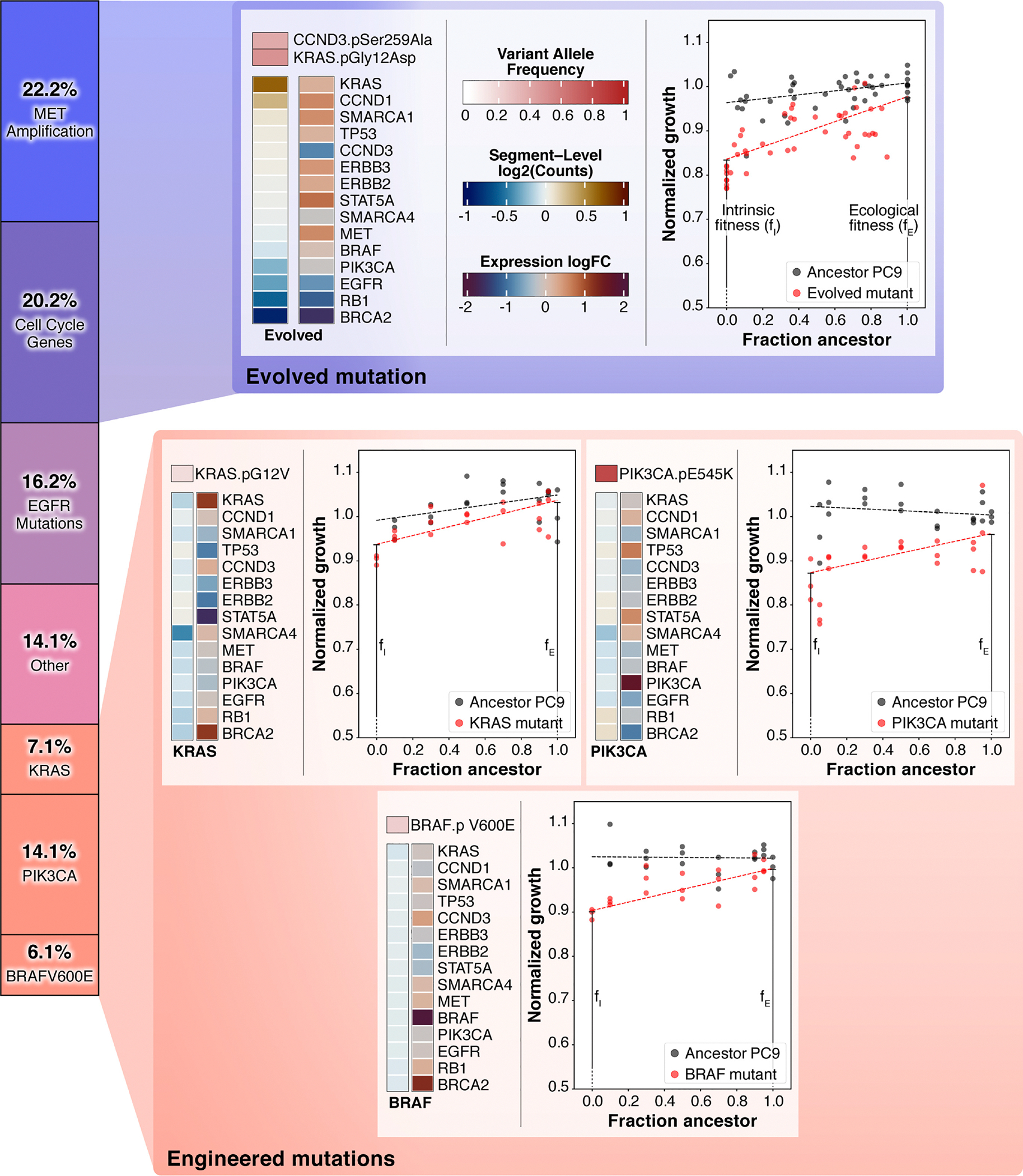
The evolution of resistance remains one of the primary challenges for modern medicine, from infectious diseases to cancers. Many of these resistance-conferring mutations often carry a substantial fitness cost in the absence of treatment. As a result, we would expect these mutants to undergo purifying selection and be rapidly driven to extinction. Nevertheless, preexisting resistance is frequently observed from drug-resistant malaria to targeted cancer therapies in non-small-cell lung cancer (NSCLC) and melanoma. Solutions to this apparent paradox have taken several forms, from spatial rescue to simple mutation supply arguments. Recently, in an evolved resistant NSCLC cell line, we found that frequency-dependent ecological interactions between ancestor and resistant mutant ameliorate the cost of resistance in the absence of treatment. Here, we hypothesize that frequency-dependent ecological interactions in general play a major role in the prevalence of preexisting resistance. We combine numerical simulations with robust analytical approximations to provide a rigorous mathematical framework for studying the effects of frequency-dependent ecological interactions on the evolutionary dynamics of preexisting resistance. First, we find that ecological interactions significantly expand the parameter regime under which we expect to observe preexisting resistance. Next, even when positive ecological interactions between mutants and ancestors are rare, these resistant clones provide the primary mode of evolved resistance because even weak positive interaction leads to significantly longer extinction times. We then find that even in the case where mutation supply alone is sufficient to predict preexisting resistance, frequency-dependent ecological forces still contribute a strong evolutionary pressure that selects for increasingly positive ecological effects (negative frequency-dependent selection). Finally, we genetically engineer several of the most common clinically observed resistance mechanisms to targeted therapies in NSCLC, a treatment notorious for preexisting resistance. We find that each engineered mutant displays a positive ecological interaction with their ancestor. As a whole, these results suggest that frequency-dependent ecological effects can play a crucial role in shaping the evolutionary dynamics of preexisting resistance.
Figures
Update of
-
Frequency-dependent ecological interactions increase the prevalence, and shape the distribution, of pre-existing drug resistance.bioRxiv [Preprint]. 2024 Mar 11:2023.03.16.533001. doi: 10.1101/2023.03.16.533001. bioRxiv. 2024. Update in: PRX Life. 2024 Apr-Jun;2(2):023010. doi: 10.1103/prxlife.2.023010. PMID: 36993678 Free PMC article. Updated. Preprint.
Similar articles
-
Frequency-dependent ecological interactions increase the prevalence, and shape the distribution, of pre-existing drug resistance.bioRxiv [Preprint]. 2024 Mar 11:2023.03.16.533001. doi: 10.1101/2023.03.16.533001. bioRxiv. 2024. Update in: PRX Life. 2024 Apr-Jun;2(2):023010. doi: 10.1103/prxlife.2.023010. PMID: 36993678 Free PMC article. Updated. Preprint.
-
Prescription of Controlled Substances: Benefits and Risks.2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 30726003 Free Books & Documents.
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2021 Apr 19;4(4):CD011535. doi: 10.1002/14651858.CD011535.pub4. Cochrane Database Syst Rev. 2021. Update in: Cochrane Database Syst Rev. 2022 May 23;5:CD011535. doi: 10.1002/14651858.CD011535.pub5. PMID: 33871055 Free PMC article. Updated.
Cited by
-
Deciphering and steering population-level response under spatial drug heterogeneity on microhabitat structures.bioRxiv [Preprint]. 2025 Jun 17:2025.02.13.638200. doi: 10.1101/2025.02.13.638200. bioRxiv. 2025. PMID: 40027692 Free PMC article. Preprint.
-
Linking spatial drug heterogeneity to microbial growth dynamics in theory and experiment.bioRxiv [Preprint]. 2024 Nov 28:2024.11.21.624783. doi: 10.1101/2024.11.21.624783. bioRxiv. 2024. PMID: 39605592 Free PMC article. Preprint.
-
Using spatial statistics to infer game-theoretic interactions in an agent-based model of cancer cells.bioRxiv [Preprint]. 2025 Jul 15:2025.07.09.664005. doi: 10.1101/2025.07.09.664005. bioRxiv. 2025. PMID: 40791473 Free PMC article. Preprint.
References
-
- Darby EM et al. , Molecular mechanisms of antibiotic resistance revisited, Nat. Rev. Microbiol. 21, 280 (2023). - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials