

Tau uptake by human neurons depends on receptor LRP1 and kinase LRRK2
- PMID: 40790356
- PMCID: PMC12436637
- DOI: 10.1038/s44318-025-00514-0
Tau uptake by human neurons depends on receptor LRP1 and kinase LRRK2
Abstract
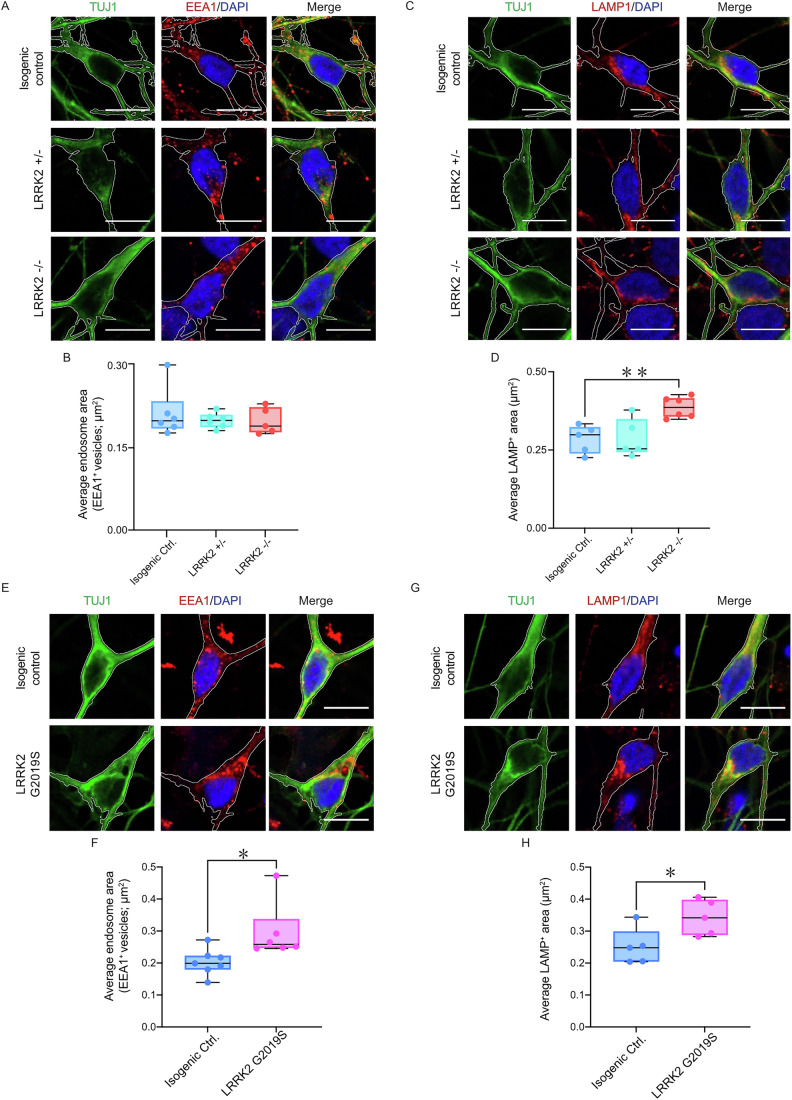
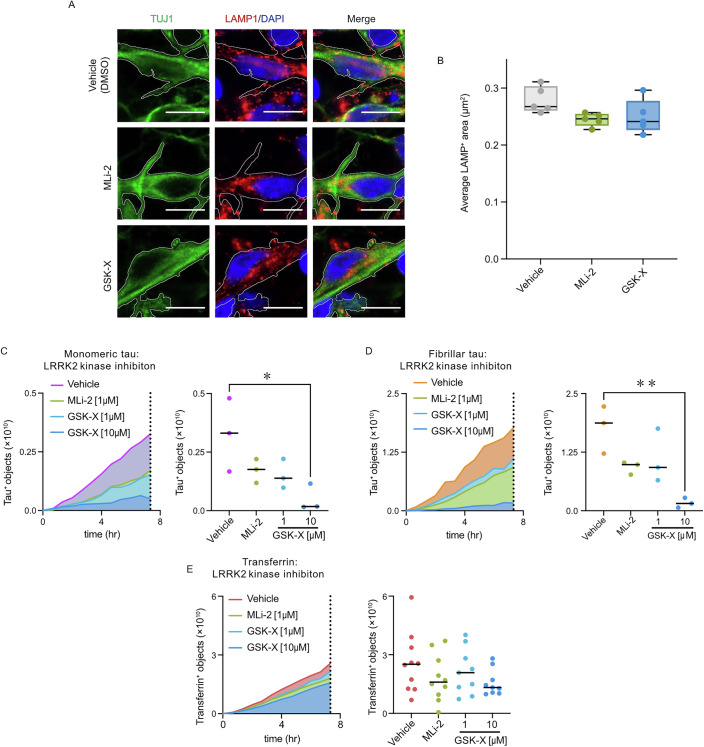
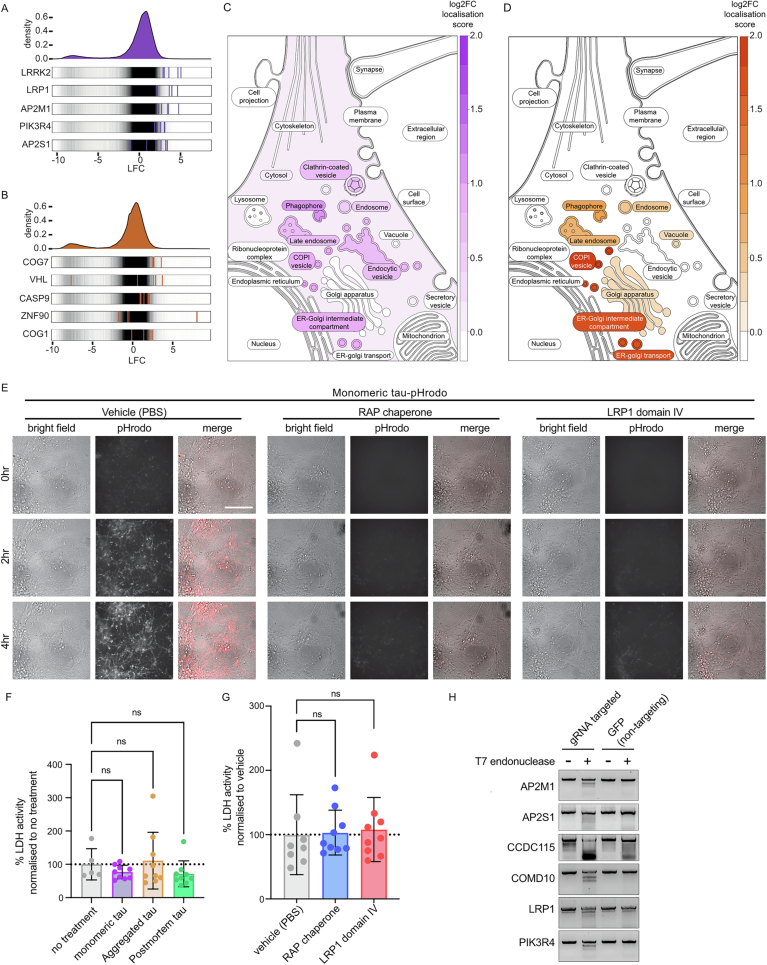
Extracellular release and uptake of pathogenic forms of the microtubule-associated protein tau contribute to the pathogenesis of several neurodegenerative diseases, including Alzheimer's disease. Defining the cellular mechanisms and pathways for tau entry to human neurons is essential to understanding tauopathy pathogenesis and enabling the rational design of disease-modifying therapeutics. Here, whole-genome, loss-of-function CRISPR screens in human iPSC-derived excitatory neurons, the major neuronal cell type affected in these diseases, provide insights into the different cellular pathways for uptake of extracellular monomeric and fibrillar tau. Monomeric and fibrillar tau are both taken up by human neurons by receptor-mediated endocytosis, but involve different routes of entry at the neuronal surface: the low-density lipoprotein LRP1 is the primary receptor for monomeric tau, but contributes less to fibrillar tau entry. Similarly, endocytosis of monomeric tau is dependent on the familial Parkinson's disease gene LRRK2, but not required for endocytosis of fibrillar tau. These findings implicate LRP1 and LRRK2 in the pathogenesis of tauopathies and Parkinson's disease, and identify LRRK2 as a potential therapeutic target for altering progression of these diseases.
Keywords: Alzheimer’s Disease; CRISPR; Functional Genomics; Human iPSCs; Parkinson’s Disease.
© 2025. The Author(s).
Conflict of interest statement
Disclosure and competing interests statement. Open Targets is a public–private partnership between non-profit research institutions and the pharmaceutical industry. FJL is a founder and holds equity in Gen2 Neuroscience and Talisman Therapeutics.
Figures











References
-
- Almende BV, Thieurmel B, Robert T (2022) visNetwork: network visualization using ‘vis.js’ library. R package version 2.0.9.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
