

This is a preprint.
Hierarchical Neural Circuit Theory of Normalization and Inter-areal Communication
- PMID: 40791322
- PMCID: PMC12338607
- DOI: 10.1101/2025.07.15.664935
Hierarchical Neural Circuit Theory of Normalization and Inter-areal Communication
Abstract
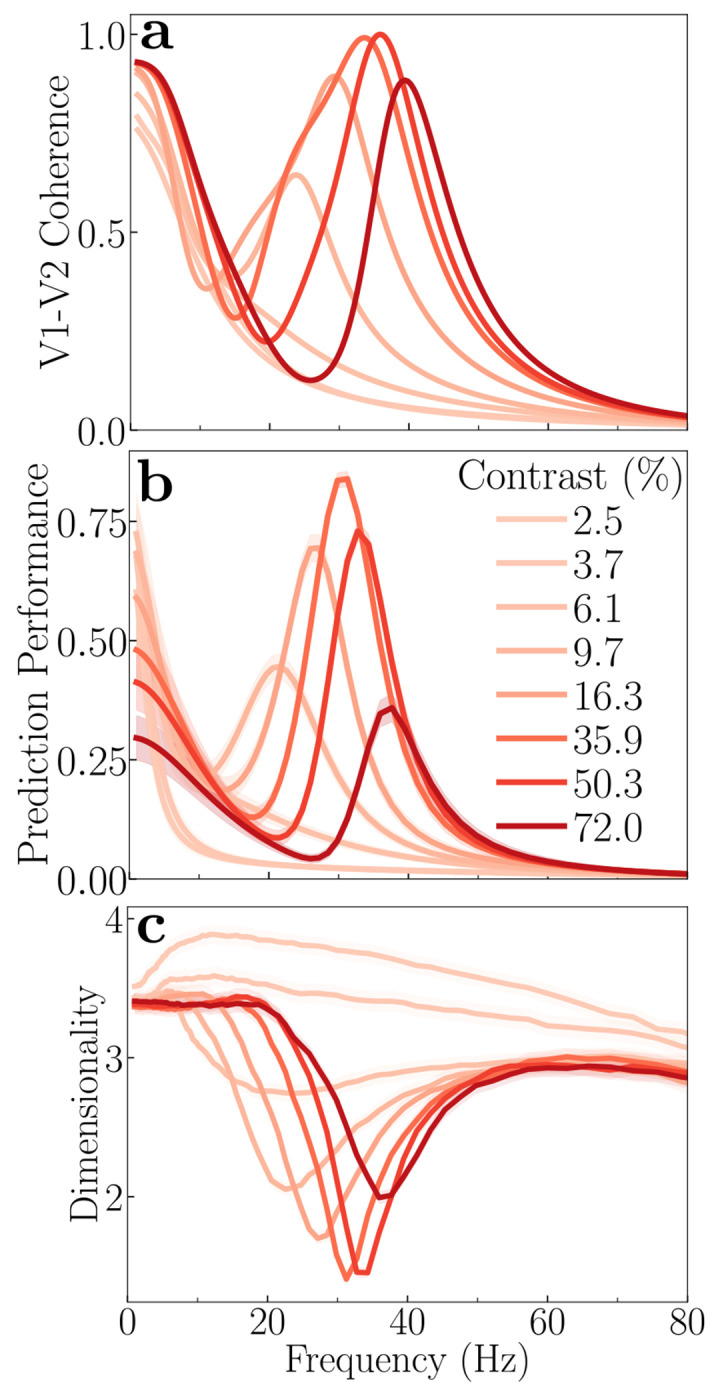
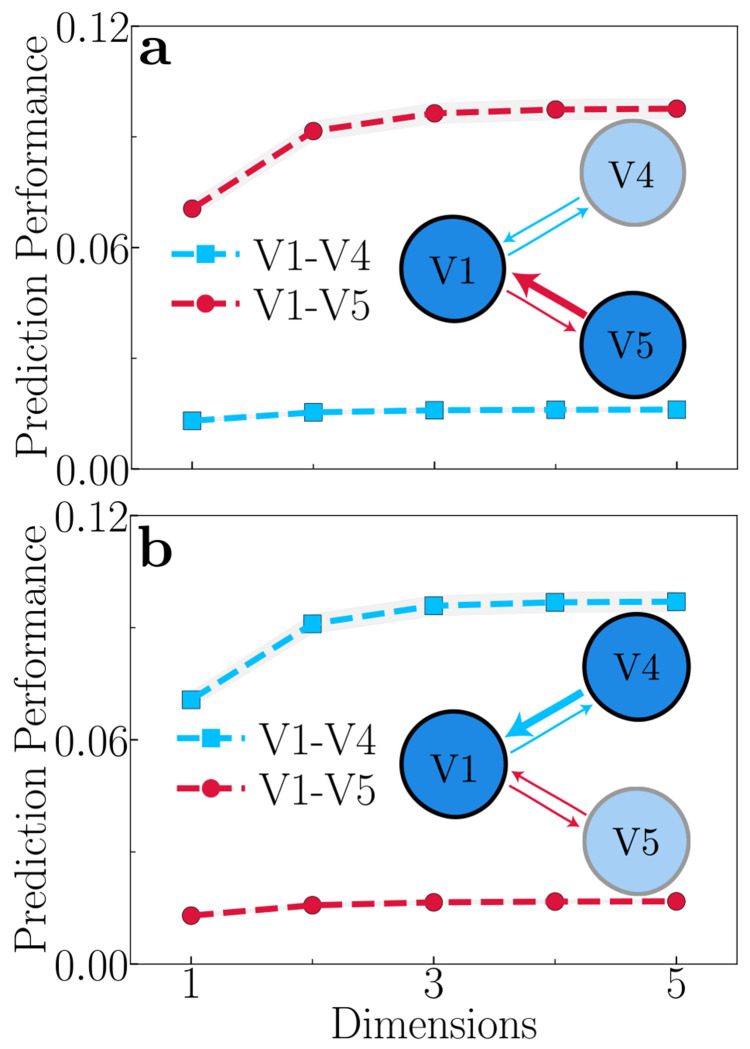
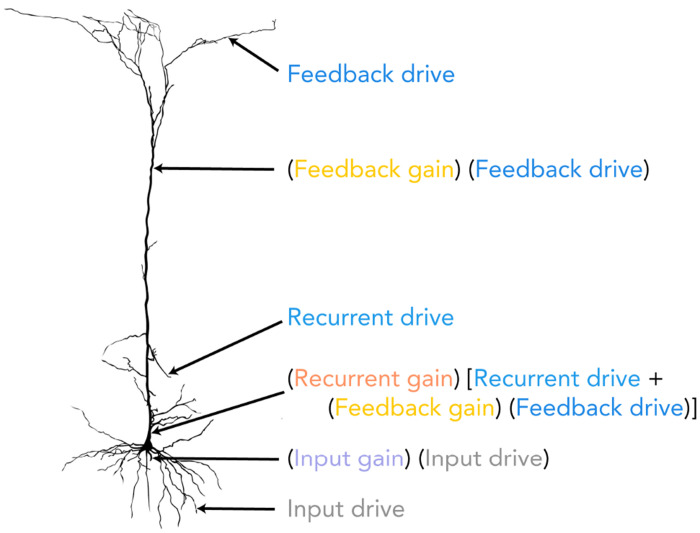
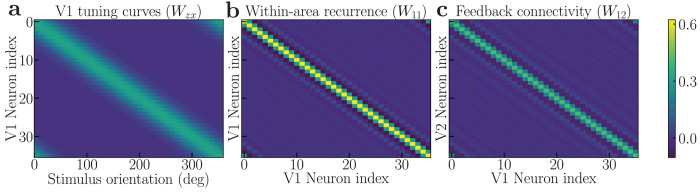
The primate brain exhibits a hierarchical, modular architecture with conserved microcircuits executing canonical computations across reciprocally connected cortical areas. Though feedback connections are ubiquitous, their functions remain largely unknown. To investigate the role of feedback, we present a hierarchical neural circuit theory with feedback connections that dynamically implements divisive normalization across its hierarchy. In a two-stage instantiation (V1 ↔ V2), increasing feedback from V2 to V1 amplifies responses in both areas, more so in the higher cortical area, consistent with experiments. We analytically derive power spectra (V1) and coherence spectra (V1-V2), and validate them against experimental observations: peaks in both spectra shift to higher frequencies with increased stimulus contrast, and power decays as 1/f 4 at high frequencies (f). The theory further predicts distinctive spectral signatures of feedback and input gain modulation. Crucially, the theory offers a unified view of inter-areal communication, with emergent features commensurate with empirical observations of both communication subspaces and inter-areal coherence. It admits a low-dimensional communication subspace, where inter-areal communication is lower-dimensional than within-area communication, and frequency bands characterized by high inter-areal coherence. It further predicts that: i) increasing feedback strength enhances inter-areal communication and diminishes within-area communication, without altering the subspace dimensionality; ii) high-coherence frequencies are characterized by stronger communication (ability to estimate neural activity in one brain area from neural activity in another brain area) and reduced subspace dimensionality. Finally, a three-area (V1 ↔ V4 and V1 ↔ V5) instantiation of the theory demonstrates that differential feedback from higher to lower cortical areas dictates their dynamic functional connectivity. Altogether, our theory provides a robust and analytically tractable framework for generating experimentally-testable predictions about normalization, inter-areal communication, and functional connectivity.
Figures



Similar articles
-
Prescription of Controlled Substances: Benefits and Risks.2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 30726003 Free Books & Documents.
-
Aspects of Genetic Diversity, Host Specificity and Public Health Significance of Single-Celled Intestinal Parasites Commonly Observed in Humans and Mostly Referred to as 'Non-Pathogenic'.APMIS. 2025 Sep;133(9):e70036. doi: 10.1111/apm.70036. APMIS. 2025. PMID: 40923351 Free PMC article. Review.
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
-
"There's Only So Much the School Can Change About Itself … Before You Need to Change Something About Yourself"-a Qualitative Analysis of the Experiences of Neurodivergent Student Teachers.Autism Adulthood. 2025 Aug 11;7(4):435-446. doi: 10.1089/aut.2024.0047. eCollection 2025 Aug. Autism Adulthood. 2025. PMID: 40933689
-
Audit and feedback: effects on professional practice.Cochrane Database Syst Rev. 2025 Mar 25;3(3):CD000259. doi: 10.1002/14651858.CD000259.pub4. Cochrane Database Syst Rev. 2025. PMID: 40130784
References
-
- Heeger David J. “Normalization of cell responses in cat striate cortex”. In: Visual neuroscience 9.2 (1992), pp. 181–197. - PubMed
-
- Salin Paul-Antoine and Bullier Jean. “Corticocortical connections in the visual system: structure and function”. In: Physiological reviews 75.1 (1995), pp. 107–154. - PubMed
-
- Shipp Stewart and Zeki Semir. “The organization of connections between areas V5 and V2 in macaque monkey visual cortex”. In: European Journal of Neuroscience 1.4 (1989), pp. 333– 354. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources