

Principles of cotranslational mitochondrial protein import
- PMID: 40795856
- PMCID: PMC12396113
- DOI: 10.1016/j.cell.2025.07.021
Principles of cotranslational mitochondrial protein import
Abstract
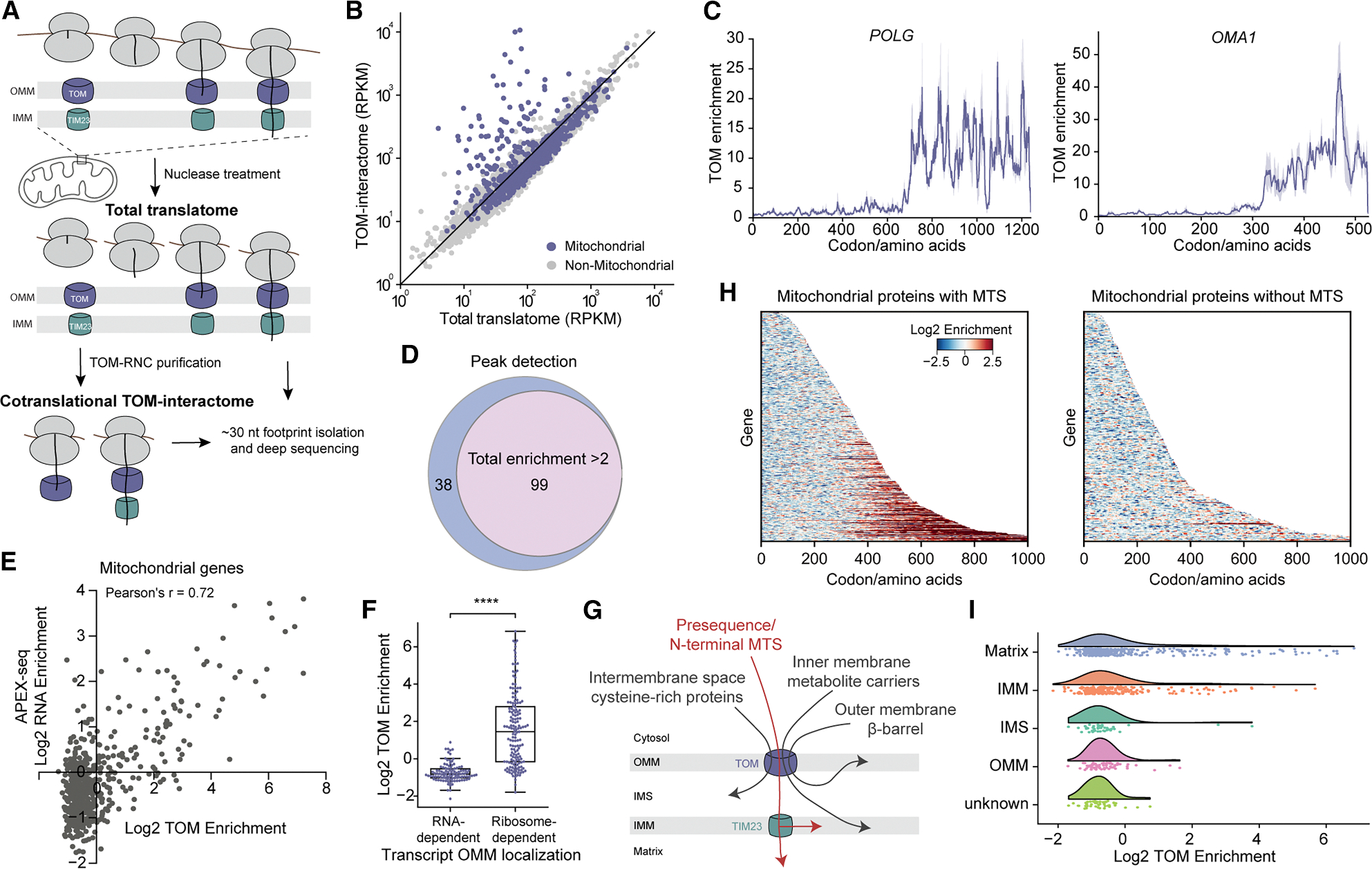
Nearly all mitochondrial proteins are translated on cytosolic ribosomes. How these proteins are subsequently delivered to mitochondria remains poorly understood. Using selective ribosome profiling, we show that nearly 20% of mitochondrial proteins can be imported cotranslationally in human cells. Cotranslational import requires an N-terminal presequence on the nascent protein and contributes to localized translation at the mitochondrial surface. This pathway does not favor membrane proteins but instead prioritizes large, multi-domain, topologically complex proteins, whose import efficiency is enhanced when targeted cotranslationally. In contrast to the early onset of cotranslational protein targeting to the endoplasmic reticulum (ER), the presequence on mitochondrial proteins is inhibited from initiating targeting early during translation until a large globular domain emerges from the ribosome. Our findings reveal a multi-layered protein sorting strategy that controls the timing and specificity of mitochondrial protein targeting.
Keywords: NAC; TOM complex; cotranslational protein import; localized translation; mitochondria; mitochondrial targeting sequence; nascent polypeptide-associated complex; protein folding; protein targeting; ribosome profiling.
Copyright © 2025 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests Authors declare no competing interests.
Figures



References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
