

This is a preprint.
Defined human tri-lineage brain microtissues
- PMID: 40799568
- PMCID: PMC12340862
- DOI: 10.1101/2025.08.05.668605
Defined human tri-lineage brain microtissues
Abstract
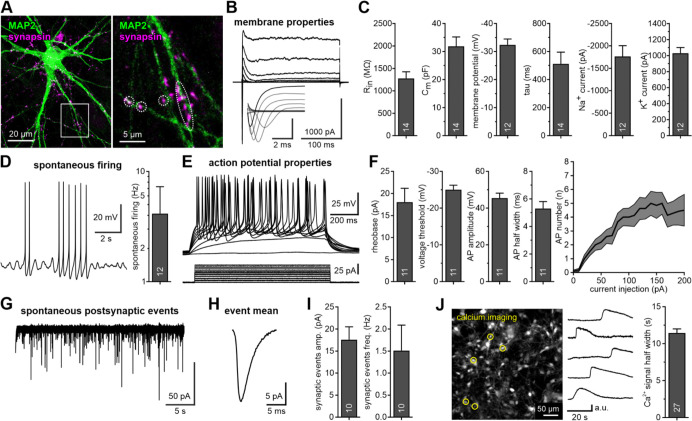
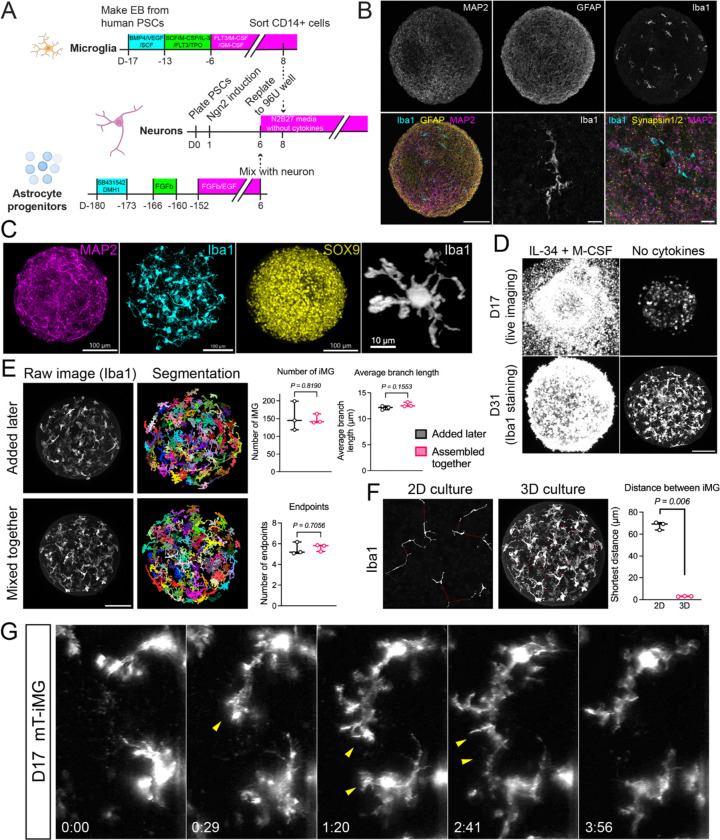
Microglia are the immune cells of the central nervous system and are thought to be key players in both physiological and disease conditions. Several microglial features are poorly conserved between mice and human, such as the function of the neurodegeneration-associated immune receptor Trem2. Induced pluripotent stem cell (iPSC)-derived microglia offer a powerful opportunity to generate and study human microglia. However, human iPSC-derived microglia often exhibit activated phenotypes in vitro, and assessing their impact on other brain cell types remains challenging due to limitations in current co-culture systems. Here, we developed fully defined brain microtissues, composed of human iPSC-derived neurons, astrocytes, and microglia, co-cultured in 2D or 3D formats. Our microtissues are stable and self-sufficient over time, requiring no exogenous cytokines or growth factors. All three cell types exhibit morphologies characteristic of their in vivo environment and show functional properties. Co-cultured microglia develop more homeostatic phenotypes compared to microglia exposed to exogenous cytokines. Hence, these tri-cultures provide a unique approach to investigate cell-cell interactions between brain cell types. We found that astrocytes and not neurons are sufficient for microglial survival and maturation, and that astrocyte-derived M-CSF is essential for microglial survival. Single-cell and single-nucleus RNA sequencing analyses nominated a network of reciprocal communication between cell types. Brain microtissues faithfully recapitulated pathogenic α-synuclein seeding and aggregation, suggesting their usefulness as human cell models to study not only normal but also pathological cell biological processes.
Conflict of interest statement
DECLARATION OF INTERESTS M.W. is a co-founder of Neucyte Inc., a scientific advisor for bit.bio Ltd, co-founder and scientific advisor of Lytherian Therapeutics and Theseus Therapies. E.U. has financial interest in Synapticure. M.B. declares outside interest in DEM Biopharma and Stylus Medicine. T.C.S. is a co-founder of Neucyte Inc. and serves as an SAB member for BitBio Ltd. H.A.L. is the co-founder and chief scientific officer of ND BioSciences, Epalinges, Switzerland and received funding from Merck Serono, AC Immune, UCB, and AbbVie. TWC is co-founder and SAB member of Qinotto Inc., Teal Rise Inc., and Vero Biosciences.
Figures





References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials