

doi: 10.1038/318630a0.
The Drosophila developmental gene, engrailed, encodes a sequence-specific DNA binding activity
- PMID: 4079979
- PMCID: PMC2880619
- DOI: 10.1038/318630a0
Item in Clipboard
The Drosophila developmental gene, engrailed, encodes a sequence-specific DNA binding activity
Nature.
1985.
Abstract
Plasmid expression vectors carrying either the entire engrailed coding region or a subfragment including the homoeo box, produce protein fusions having sequence-specific DNA binding activity.
Figures
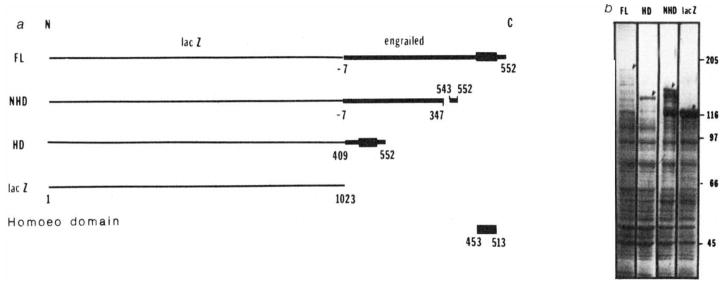
Construction of lacZ-engrailed fusions and expression in Escherichia coli. a, Gene fusions; b, polyacrylamide gel electrophoresis of bacterial extracts; FL, full-length fusion; HD, homoeo domain fusion; NHD, non-homoeo domain fusion; lacZ, β-galactosidase. Methods: a, DNA fragments derived from the engrailed locus,, were inserted in the polylinker of expression vector plasmids pUR 290, 291 or 292 (ref. 47). These different plasmids allow inserted DNA to be expressed as a C-terminal extension of β-galactosidase. In the full-length fusion protein (FL) the extension includes 7 amino acids that precede the first methionine of the engrailed protein as well as the entire engrailed protein. In the homoeo domain construct (HD), a BamHI fragment was spliced out of the FL construction, keeping only the last quarter of the engrailed coding sequence from amino acid 409 to the end. This fusion protein contains the entire homoeo domain (amino acids 453–513). Amino acids 347–542, containing the homoeo domain, have been deleted in the non-homoeo domain protein (NHD) by removing a XhoI fragment from the FL construct. In the absence of insertion the lacZ gene is expressed as the complete β-galactosidase. b, Polyacrylamide gel electrophoresis of bacterial extracts. The expression of the fusion proteins from chimaeric plasmids is under the control of the lac promoter in a lacI overproducing strain DG101 (a gift from D. Gelfand, Cetus Corporation). Bacteria in exponential growth were induced with isopropyl β-thiogalactoside at an absorbance of 0.5 (at 600 nm) and collected 2 h later. Cells were resuspended in about 0.005 culture volume of 25% sucrose, 0.2 mM EDTA, 40 mM Tris-HCI pH 7.5 and 1 mM dithiothreitol (DTT). Lysis and protein solubilization were achieved by lysozyme treatment (0.4 mg ml−1 for 1 h at 0 °C followed by addition of urea to 4 M and further incubation at 0 °C for 1 h). After centrifugation at 20,000 r.p.m. for 1 h the urea was removed from the supernatant by dialysis first against 10 mM Tris-HCI pH 7.5, 25 mM NaCl, 1 mM EDTA, 0.1% Triton X-100, 1 mM DTT, 10% glycerol, 1 mM phenylmethylsulphonyl fluoride, and 0.1 mM benzamidine containing 2 M urea and subsequently against the same buffer without urea. Glycerol was added to the extracts to a final concentration of 50% and they were stored at −20 °C. When analysed by SDS-gel electrophoresis the fusion proteins are seen as abundant high relative molecular mass proteins (arrowheads). The larger fusion protein is present in lower amounts and we detect numerous minor bands that are presumed to result from degradation of the fusion proteins.
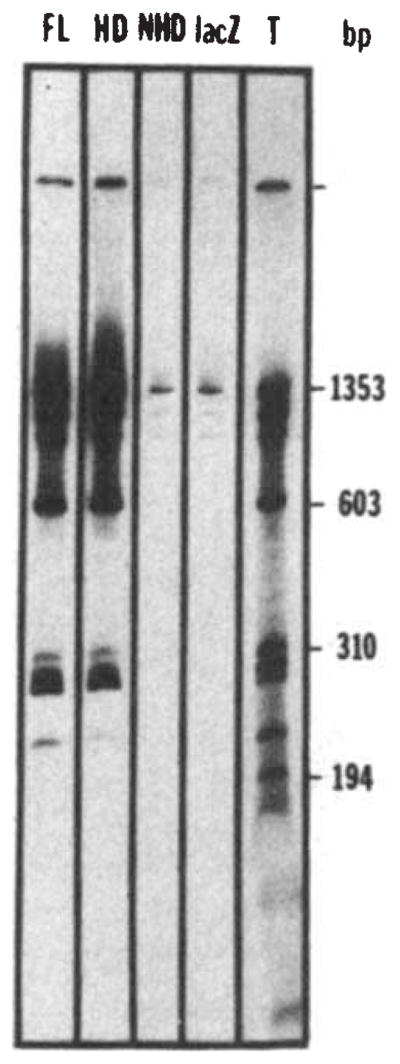
Nonspecific binding of the fusion proteins to DNA. Bacteriophage Φ×174 DNA was cleaved by HaeIII and end-labelled using T4 polymerase. The labelled DNA (about 30 ng) was incubated for 30 min at 0 °C in 25 μ1 of binding buffer (BB) (50 mM NaCl, 20 mM Tris pH 7.6, 0.25 mM EDTA, 1 mM DTT, 10% glycerol) in the presence of a bacterial extract containing the fusion protein (about 10 μg of total protein extract, see Fig. 1 legend). The complexes formed between DNA and the fusion proteins were then immunoprecipitated by 30 min further incubation at 0 °C with an anti-β-galactosidase monoclonal antibody (provided by Tom Mason and Judy Partaledis, courtesy of Mike Hall) preadsorbed on cross-linked Staphylococcus (Pansorbin, Calbiochem). The pellet was washed twice in BB, phenol extracted and the DNA ethanol precipitated before polyacrylamide gel electrophoresis and autoradiography. The autoradiogram shows the results of precipitations performed in the presence of extracts containing the full-length fusion (FL), the homoeo domain fusion (HD), the non-homoeo domain fusion (NHD) or β-galactosidase (lacZ). Lane T shows the labelled digest before immunoprecipitation, representing 25% of the counts added to the incubation mixture in the immunoprecipitation experiments.
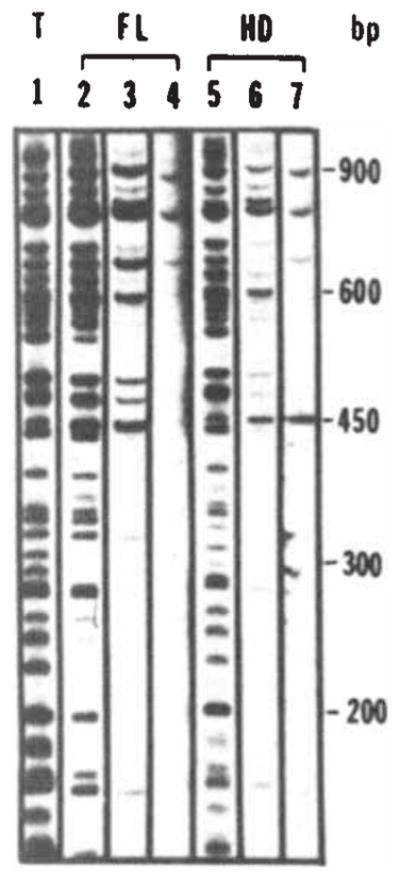
Sequence-specific DNA interaction of the fusion protein with bacteriophage λ DNA fragments. Bacteriophage λ DNA was restricted with Sau3A and labelled using T4 polymerase. Binding assays were performed as described in Fig. 2 legend with the addition of carrier DNA as indicated. Fragments were separated on a 5% polyacrylamide gel. Lane 1 shows the total digest. The amount of DNA loaded in lane 1 is one-quarter of the input amount used in the binding assays (10ng; 0.4 μg ml−1). Lanes 2, 3 and 4 show immunoprecipitates obtained following addition of the full-length fusion extract in the presence of 0, 4 and 40 μg ml−1 of carrier DNA (fragmented calf thymus DNA), respectively. Lanes 5, 6 and 7 show the results of a similar experiment using the homoeo domain fusion extract. Lanes 4 and 7 were exposed four times longer because of the reduced recovery of bands at high levels of carrier.
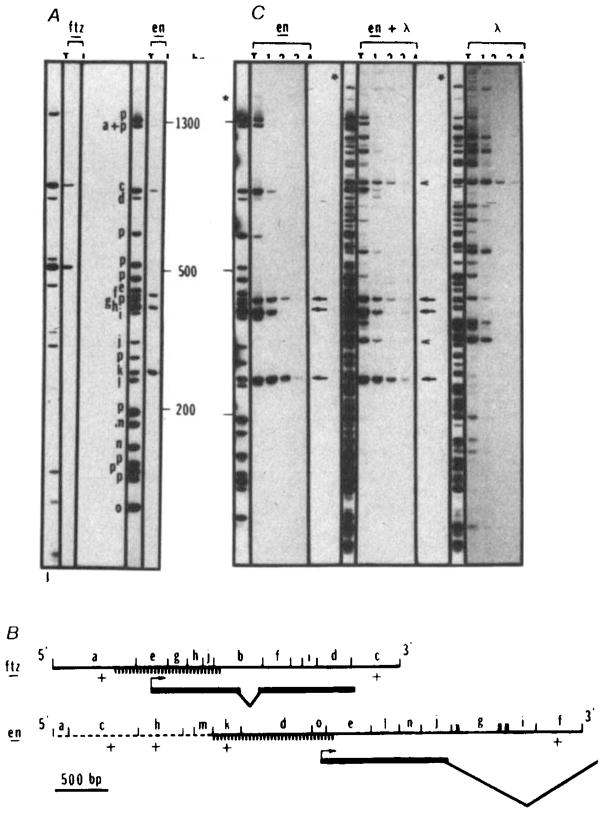
Sequence-specific interaction of the homoeo domain fusion protein with restriction fragments of cloned fushi tarazu and engrailed sequences. The fushi tarazu clone (p6-3, derived from clone pDmA439, a gift from Matt Scott) contains 900 bp of upstream sequence, the entire coding sequence and 800 bp of downstream sequence. The engrailed clone (p615) contains 2.3 kb of upstream sequence, the complete first exon and most of the first intron, (panel B). Restriction fragments of p6-3, p615 or λ DNA were end-labelled and tested for fusion protein binding as described in the legend for Fig. 2, except that a higher salt concentration (170 mM NaCl) was used to diminish nonspecific interactions. Fragments were separated on a 5% acrylamide gel. In the example shown in A, the fushi tarazu plasmid (ftz) DNA was digested with RsaI and the engrailed plasmid (en) was digested with HinfI and ClaI. Lanes marked T show the total fragment pattern and lanes marked I show the fragments recovered in the immunoprecipitate. For both plasmids the amount of DNA loaded in T is one-quarter of the amount subjected to immunoprecipitation and displayed in I. Bands derived totally from plasmid sequences are identified with a ‘p’. Bands derived at least in part from insert sequences are designated as a to j (ftz) or a to o (en) and their positions within the cloned sequence are indicated in B. Similar experiments using AluI, HaeIII, BstNI, DdeI or HpaII to digest these plasmids gave comparable results (not shown). B shows a map of the inserts in p6-3 (ftz) and p615 (en). The orientation of the coding region within the insert is indicated. Except for the dashed portion of the engrailed insert, the sequence is known (refs , , and J. Kassis, D. K. Wright and C.D., unpublished). The positions of restriction enzyme sites (RsaI sites in ftz and HinfI and ClaI sites in en) are indicated and all fragments detectable in the separation shown in panel A are lettered. Those fragments detected as bound by the homoeo domain fusion protein are indicated as +. Because they were not detected, a few small restriction fragments (unlettered) could not be scored in the binding experiment (A) but analysis of these DNAs cleaved with other restriction enzymes failed to detect additional binding sites. Exons of the transcribed region are indicated with a bold line and introns with sloping lines. The hatched region indicates the position of a subfragment that was purified and used for additional binding studies (Fig. 5). Note that the region responsible for binding of ftz, fragment a, was further localized by analysis of AluI, HaeIII, BstNI, DdeI, HpaII and HinfI digests that defined only one site; this site corresponds to that mapped in more detail by analysis of the purified subfragment (Fig. 5). C Compares the efficiency of homoeo domain fusion protein binding to engrailed or λ sites. The binding assays were performed as described in A except that increasing amounts of unlabelled DNA (HinfI-restricted p615) was added as competitor. Lanes T represent 10% of the counts of the total digest submitted to immunoprecipitation in lanes 1, 2, 3 and 4. In lane 1, no unlabelled DNA was added while in lanes 2, 3 and 4, 30 ng, 250 ng and 4 μg, respectively, were added, en lanes: plasmid p615 was restricted with ClaI and HinfI labelled and submitted to immunoprecipitation. Lanes 1, 2, 3 and 4 represent immunoprecipitated DNA from ~4 ng of labelled DNA (~ 1 fmol). Obviously the four bands retained in A behave differently when unlabelled competitor DNA is added. Three bands (arrows) corresponding to fragments f, k and h in A, are detected in the immunoprecipitate in the presence of 1,000-fold excess of unlabelled DNA. λ lanes: λ DNA was restricted with ClaI and HinfI, labelled and processed under the same conditions as for en DNA. An equimolar amount of DNA (1 fmol) was used in this experiment (20 ng). Two bands were retained at the highest stringency (arrowheads). en + λ lanes: T is an equimolar mixture of en and λ DNA restricted with ClaI and HinfI. The bands that are retained are the same as those seen in the experiments testing en and λ DNAs individually. Five bands are retained in the presence of the highest concentration of competitor (arrows and arrowheads).
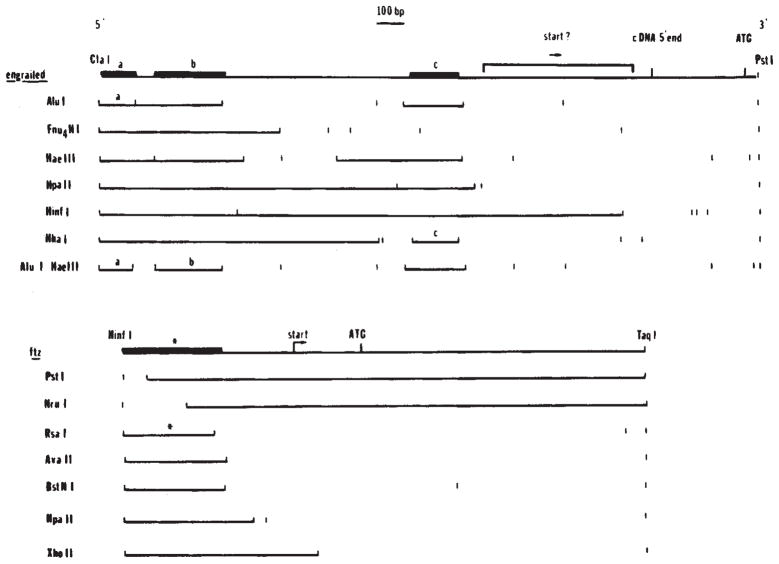
Localization of binding sites in the 5′ regions of engrailed and fushi tarazu. In digests of whole plasmid DNA we identified fragments that were bound by the homoeo domain fusion protein (Fig. 4). To localize more precisely the binding sites immediately upstream of the engrailed and fushi tarazu coding regions, we purified a 1,180-bp ClaI-PstI fragment from the engrailed clone and a 939-bp HinfI-TaqI fragment from the fushi tarazu clone (Fig. 4B). These purified fragments were digested with additional enzymes as indicated and the binding of subfragments assayed as before (Fig. 4). In each of the restriction digests shown here the binding fragments are indicated as solid lines. Although the overlap of binding sites: a ClaI-AluI fragment (a, the binding regions, this could be misleading if the binding sites were complex. Therefore, in the summary diagrams above the restriction patterns, we have indicated the precision of the localization of the binding sites based on the minimal fragment showing binding (bold line). For engrailed, three fragments were identified as containing binding sites: a, ClaI-AluI fragment (a, 67 bp), a HaeIII-AluI fragment (b, 100 bp) and a HhaI fragment (c, 88 bp). For ftz a single site was localized to a 165-bp HinfI-RsaI fragment(*).
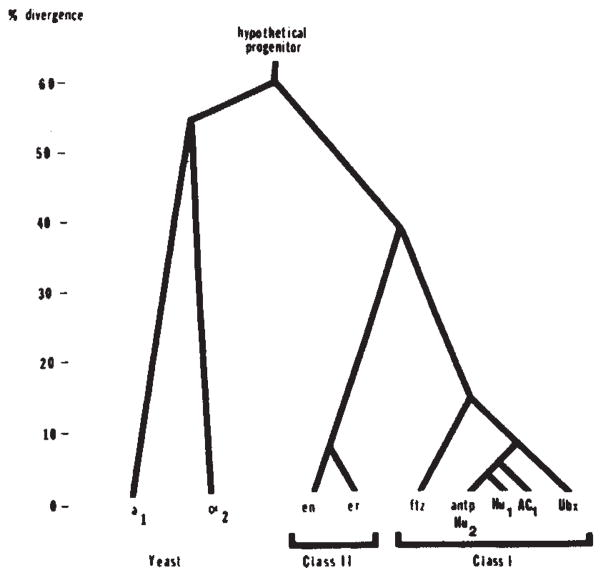
Family tree of relatedness of homoeo domains. Pairwise comparisons of the protein sequences of the homoeo domains encoded by Drosophila genes (Antp, Ubx, ftz, en and er), yeast genes (a1 and α2) and sequences isolated by homology from humans (Hu1 and Hu2) and frogs (AC1) were used to score divergence. The comparison was confined to residues 29-58 because this region is well conserved among all of these sequences. To produce homology scores we gave one point for amino-acid identity and half a point for similarity. The data are assembled into a family tree by indicating the divergence of each sequence that approximates all the pairwise comparisons. The sequence data are taken from Levine et al (Hu1 and Hu2), Scott and Weiner (Ubx and ftz), McGinnis et a1 (Antp), Poole et al. (en and er), Carraso et al. (AC1), and Tatchell et al. (α2 and a1). Note that the discrepancies in the published sequence, for Ubx are significant in comparison to the divergence among class I homoeo domains.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
