

The fluctuations of alpha power: Bimodalities, connectivity, and neural mass models
- PMID: 40800890
- PMCID: PMC12330861
- DOI: 10.1162/IMAG.a.64
The fluctuations of alpha power: Bimodalities, connectivity, and neural mass models
Abstract
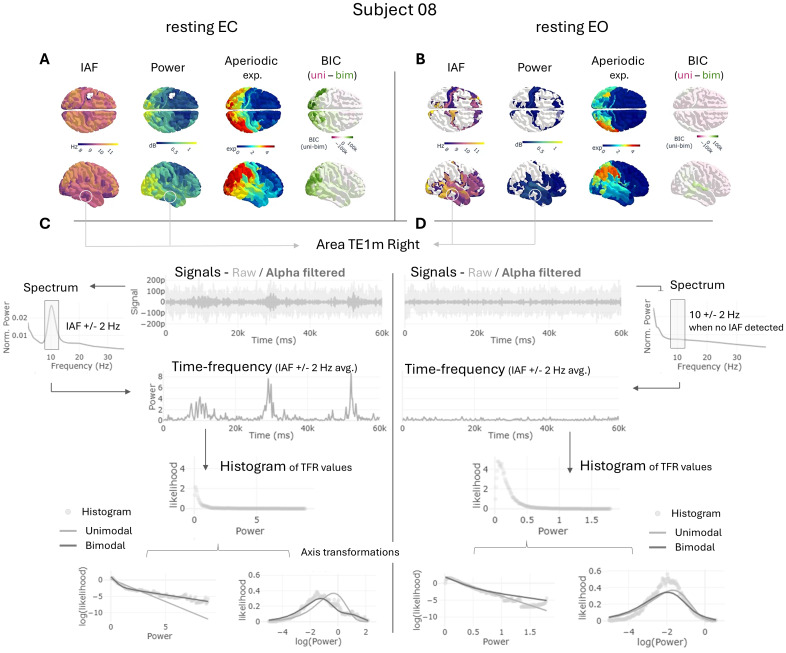
The alpha rhythm is a hallmark of electrophysiological resting-state brain activity, that serves as a biomarker in health and disease. Alpha power is far from uniform over time, exhibiting dynamic fluctuations. The likelihood of those power values can be captured by a decreasing exponential function, which in certain cases becomes bimodal. While alpha rhythm is usually evaluated through the averaged power spectra across entire recordings, its dynamic fluctuations have received less attention. In this study, we investigate the dynamic nature of alpha power, its relationship with functional connectivity (FC) within the default mode network (DMN), and the ability of the Jansen-Rit (JR) neural mass model to replicate these fluctuations. Using MRI and MEG data from 42 participants in resting state with eyes-closed and eyes-open, we evaluated the shape of the exponential distributions for alpha power fluctuations, and their relationship with other spectral variables as frequency, power, and the aperiodic exponent. Additionally, we assessed the temporal relationship between alpha power and FC using phase-based (ciPLV) and amplitude-based (cAEC) metrics. Finally, we employed diffusion-weighted MRI to construct brain network models incorporating JR neural masses to reproduce and characterize alpha fluctuations. Our results indicate that alpha power predominantly follows unimodal exponential distributions, with bimodalities associated to high-power in posterior regions. FC analyses revealed that ciPLV and cAEC were directly correlated with alpha power within the DMN in alpha and beta bands, whereas only theta-band ciPLV showed an inverse relationship with alpha power. JR model simulations suggested that post- supercritical fixed points better replicated alpha power fluctuations compared to limit cycle parameterizations and pre-saddle node fixed points. These results deepen our understanding of the dynamics of alpha rhythm and its intricate relationship with FC patterns, offering novel insights to refine biologically plausible brain simulations and advance computational models of neural dynamics.
Keywords: DMN; Jansen-Rit; MEG; alpha rhythm; computational model; resting state.
© 2025 The Authors. Published under a Creative Commons Attribution 4.0 International (CC BY 4.0) license.
Conflict of interest statement
The authors declare no competing financial or non-financial interests related to this work.
Figures





References
-
- Babiloni, C., Arakaki, X., Baez, S., Barry, R. J., Benussi, A., Blinowska, K., Bonanni, L., Borroni, B., Bayard, J. B., Bruno, G., Cacciotti, A., Carducci, F., Carino, J., Carpi, M., Conte, A., Cruzat, J., D’Antonio, F., Della Penna, S., Percio, C. D., … Kamondi, A. (2025). Alpha rhythm and Alzheimer’s disease: Has Hans Berger’s dream come true? Clinical Neurophysiology, 172, 33–50. 10.1016/j.clinph.2025.02.256 - DOI - PubMed
-
- Belouchrani, A.,Abed-Meraim, K.,Cardoso, J.-F., & Moulines, E. (1997). A blind source separation technique using second-order statistics. IEEE Transactions on Signal Processing, 45(2), 434–444. https://doi.org/10.1109/78.554307
-
- Berger, H. (1929). Über das elektrenkephalogramm des menschen. Archiv für Psychiatrie und Nervenkrankheiten, 87(1), 527–570. 10.1007/bf01797193 - DOI
LinkOut - more resources
Full Text Sources