

PRMT1-mediated metabolic reprogramming promotes leukemogenesis
- PMID: 40801789
- PMCID: PMC12349897
- DOI: 10.7554/eLife.105318
PRMT1-mediated metabolic reprogramming promotes leukemogenesis
Abstract
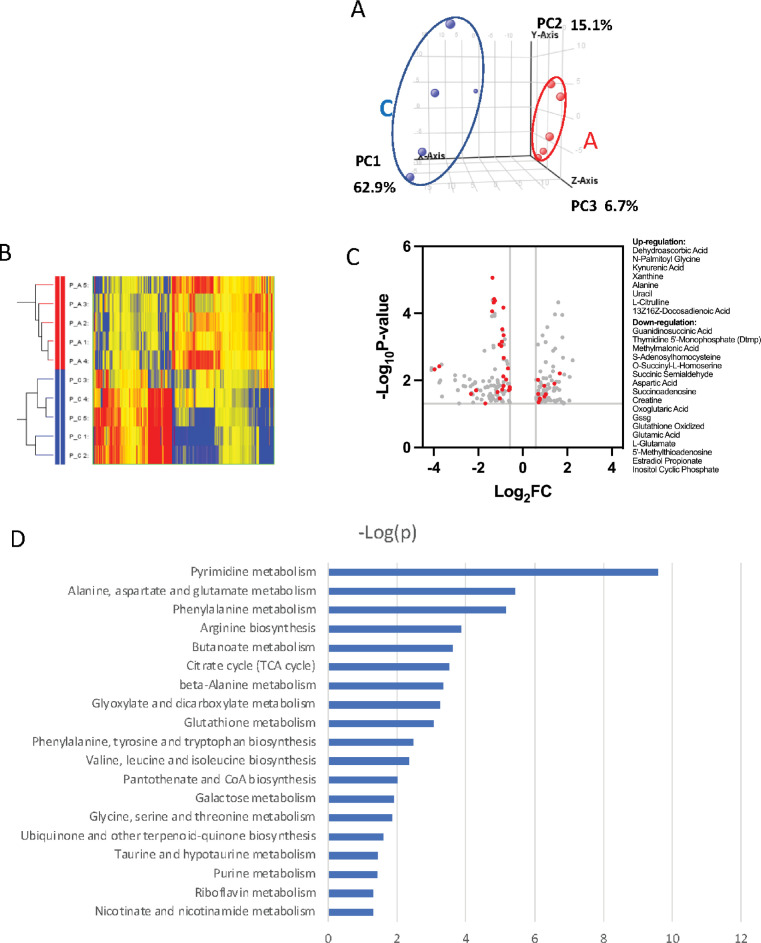
Copious expression of protein arginine methyltransferase 1 (PRMT1) is associated with poor survival in many types of cancers, including acute myeloid leukemia. We observed that a specific acute megakaryocytic leukemia (AMKL) cell line (6133) derived from RBM15-MKL1 knock-in mice exhibited heterogeneity in Prmt1 expression levels. Interestingly, only a subpopulation of 6133 cells expressing high levels of Prmt1 caused leukemia when transplanted into congenic mice. The PRMT1 inhibitor, MS023, effectively cured this PRMT1-driven leukemia. Seahorse analysis revealed that PRMT1 increased the extracellular acidification rate and decreased the oxygen consumption rate. Consistently, PRMT1 accelerated glucose consumption and led to the accumulation of lactic acid in the leukemia cells. The metabolomic analysis supported that PRMT1 stimulated the intracellular accumulation of lipids, which was further validated by fluorescence-activated cell sorting analysis with BODIPY 493/503. In line with fatty acid accumulation, PRMT1 downregulated the protein level of CPT1A, which is involved in the rate-limiting step of fatty acid oxidation. Furthermore, administering the glucose analog 2-deoxy-D-glucose delayed AMKL progression and promoted cell differentiation. Ectopic expression of Cpt1a rescued the proliferation of 6133 cells ectopically expressing PRMT1 in the glucose-minus medium. In conclusion, PRMT1 upregulates glycolysis and downregulates fatty acid oxidation to enhance the proliferation capability of AMKL cells. .
Keywords: CPT1A; PRMT1; cancer biology; cell biology; fatty acids; glycolysis; leukemia; mitochondria; mouse.
© 2025, Su et al.
Conflict of interest statement
HS, YS, HG, CS, QC, SL, AL, MG, RZ, GR, JJ, CQ, MY, CK, GZ, SB, MK, LZ, ZC, CS, SG, YC, ML, XZ No competing interests declared
Figures










Update of
- doi: 10.1101/2024.12.12.628174
- doi: 10.7554/eLife.105318.1
- doi: 10.7554/eLife.105318.2
References
-
- Avasarala S, Van Scoyk M, Karuppusamy Rathinam MK, Zerayesus S, Zhao X, Zhang W, Pergande MR, Borgia JA, DeGregori J, Port JD, Winn RA, Bikkavilli RK. PRMT1 is a novel regulator of epithelial-mesenchymal-transition in non-small cell lung cancer. The Journal of Biological Chemistry. 2015;290:13479–13489. doi: 10.1074/jbc.M114.636050. - DOI - PMC - PubMed
-
- Baldwin RM, Morettin A, Paris G, Goulet I, Côté J, Baldwin RM, Morettin A, Paris G, Goulet I, Côté J, Baldwin RM, Morettin A, Paris G, Goulet I, Côté J. Alternatively spliced protein arginine methyltransferase 1 isoform PRMT1v2 promotes the survival and invasiveness of breast cancer cells. Cell Cycle. 2012;11:4597–4612. doi: 10.4161/cc.22871. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
- IK6 BX005800/BX/BLRD VA/United States
- CX002706/U.S. Department of Veterans Affairs
- BX006321/U.S. Department of Veterans Affairs
- BX004426/U.S. Department of Veterans Affairs
- R01 HL158097/HL/NHLBI NIH HHS/United States
- R01 HL167201/HL/NHLBI NIH HHS/United States
- R01 AG082839/AG/NIA NIH HHS/United States
- BX005800/U.S. Department of Veterans Affairs
- AG082839/NH/NIH HHS/United States
- R35 GM131858/GM/NIGMS NIH HHS/United States
- HL167201/NH/NIH HHS/United States
- I01 BX006321/BX/BLRD VA/United States
- I01 CX002706/CX/CSRD VA/United States
- HL146103/NH/NIH HHS/United States
- R01 HL146103/HL/NHLBI NIH HHS/United States
- HL158097/NH/NIH HHS/United States
- R35 grant GM131858/NH/NIH HHS/United States
- R21 CA202390/CA/NCI NIH HHS/United States
- I01 BX004426/BX/BLRD VA/United States
LinkOut - more resources
Full Text Sources
