

Membrane curvature association of amphipathic helix 8 drives constitutive endocytosis of GPCRs
- PMID: 40802762
- PMCID: PMC12346279
- DOI: 10.1126/sciadv.adv1499
Membrane curvature association of amphipathic helix 8 drives constitutive endocytosis of GPCRs
Abstract
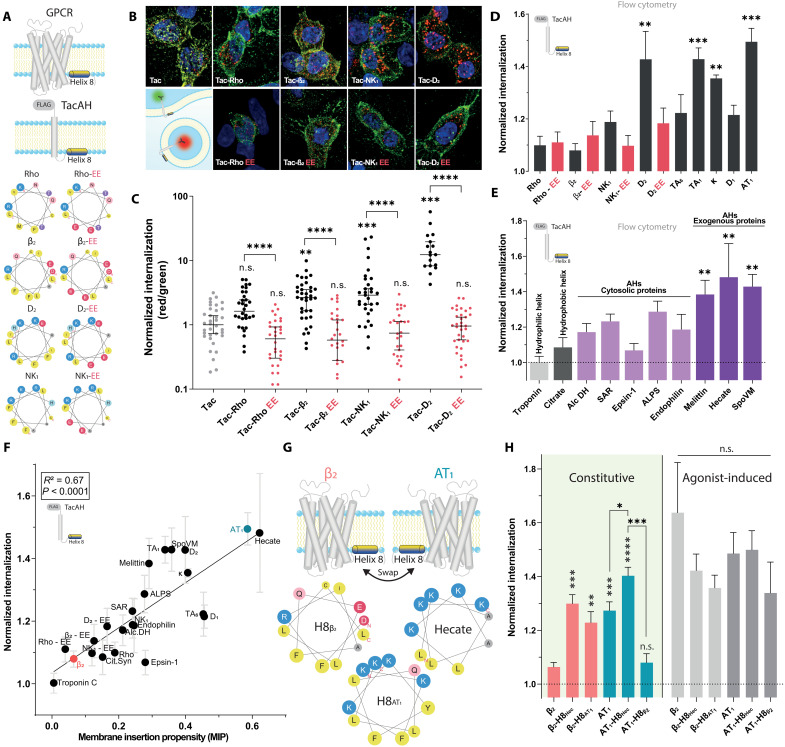
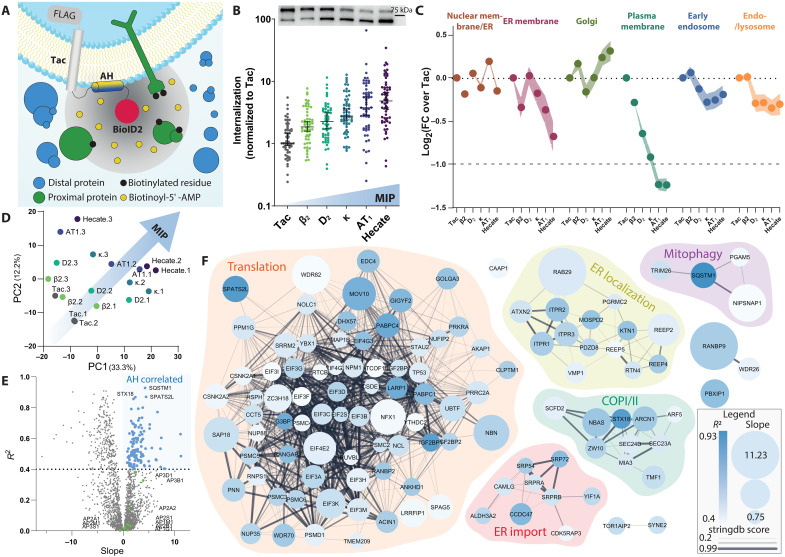
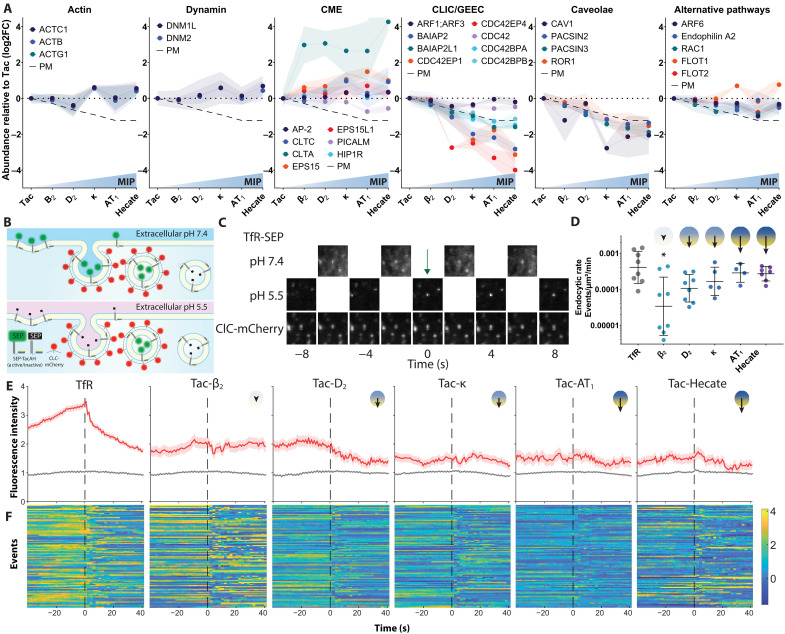
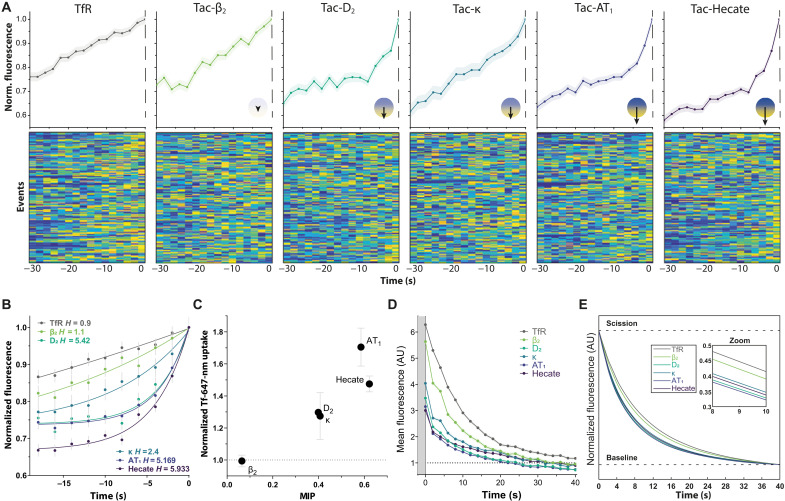
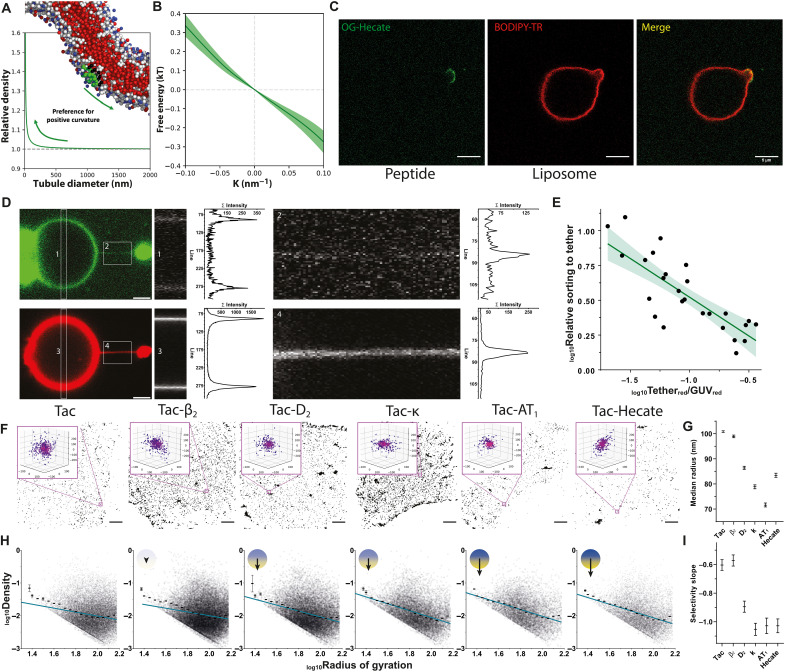
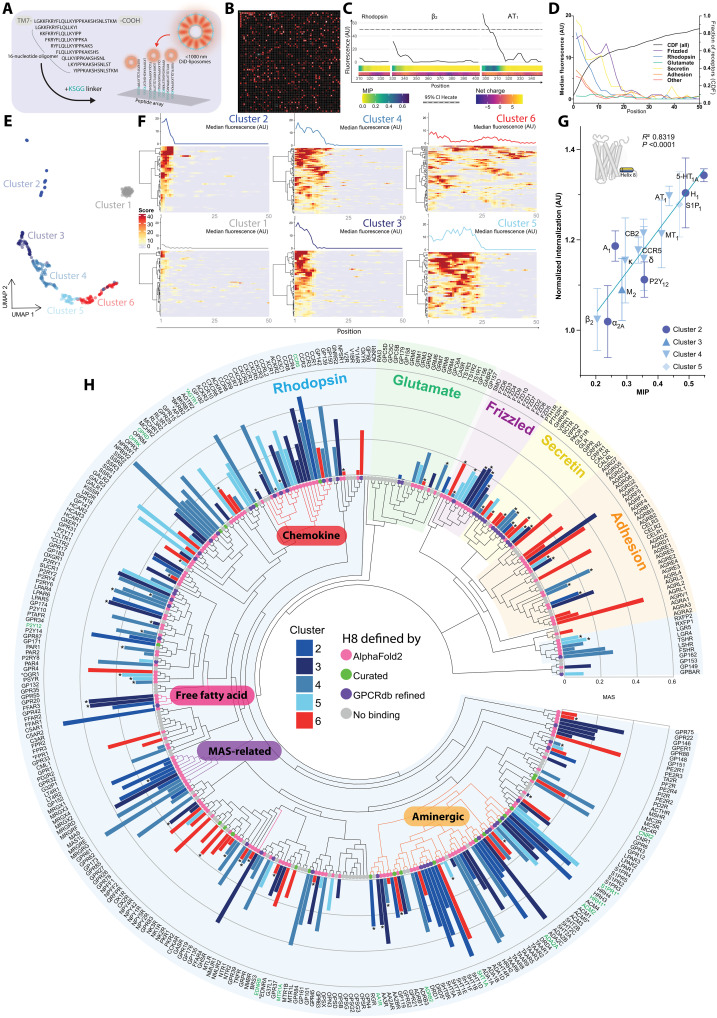
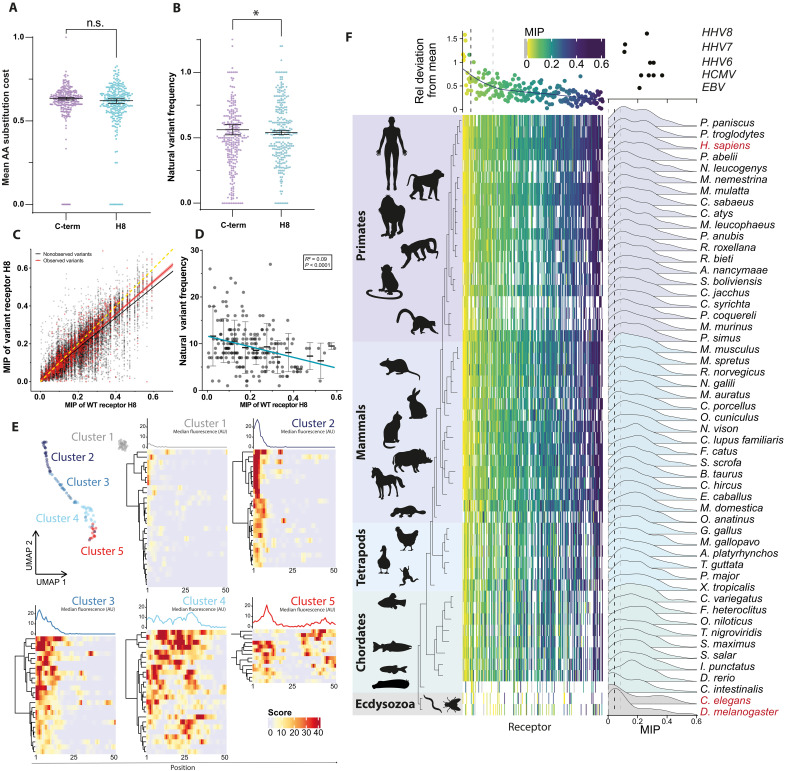
Cellular signaling relies on the activity of transmembrane receptors and their presentation on the cellular surface. Their continuous insertion in the plasma membrane is balanced by constitutive and activity-dependent internalization, which is orchestrated by adaptor proteins recognizing semispecific motifs within the receptors' intracellular regions. Here, we describe a complementary trafficking mechanism for G protein-coupled receptors (GPCRs) that is evolutionary conserved and refined. This mechanism relies on the insertion of their amphipathic helix 8 into the inner leaflet of lipid membranes, orthogonal to the transmembrane helices. These amphipathic helices dictate subcellular localization of the receptors and autonomously drive their endocytosis by cooperative assembly and association with areas of high membrane curvature. The strength of helix 8 membrane insertion propensity quantitatively predicts the rate of constitutive internalization of GPCRs. This discovery advances our understanding of membrane protein trafficking and highlights a principle of receptor-lipid interactions that may have broad implications for cellular signaling and therapeutic targeting.
Figures
Similar articles
-
HIV-1 Nef Antagonizes SERINC5 Restriction by Downregulation of SERINC5 via the Endosome/Lysosome System.J Virol. 2018 May 14;92(11):e00196-18. doi: 10.1128/JVI.00196-18. Print 2018 Jun 1. J Virol. 2018. PMID: 29514909 Free PMC article.
-
Unique hydrophobic extension of the RGS2 amphipathic helix domain imparts increased plasma membrane binding and function relative to other RGS R4/B subfamily members.J Biol Chem. 2007 Nov 9;282(45):33064-75. doi: 10.1074/jbc.M702685200. Epub 2007 Sep 11. J Biol Chem. 2007. PMID: 17848575
-
Prescription of Controlled Substances: Benefits and Risks.2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 30726003 Free Books & Documents.
-
GPCR signaling via cAMP nanodomains.Biochem J. 2025 May 13;482(10):519-33. doi: 10.1042/BCJ20253088. Biochem J. 2025. PMID: 40364615 Free PMC article. Review.
-
Deciphering complexity of GPCR signaling and modulation: implications and perspectives for drug discovery.Clin Sci (Lond). 2025 May 20;139(10):463-77. doi: 10.1042/CS20245182. Clin Sci (Lond). 2025. PMID: 40400289 Free PMC article. Review.
References
-
- McMahon H. T., Boucrot E., Molecular mechanism and physiological functions of clathrin-mediated endocytosis. Nat. Rev. Mol. Cell. Biol. 12, 517–533 (2011). - PubMed
-
- Bonifacino J. S., Traub L. M., Signals for sorting of transmembrane proteins to endosomes and lysosomes. Annu. Rev. Biochem. 72, 395–447 (2003). - PubMed
-
- Sanger A., Hirst J., Davies A. K., Robinson M. S., Adaptor protein complexes and disease at a glance. J. Cell Sci. 132, jcs222992 (2019). - PubMed
-
- Antonny B., Mechanisms of membrane curvature sensing. Annu. Rev. Biochem. 80, 101–123 (2011). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
