

Mitochondrial ROS and HIF-1α signaling mediate synaptic plasticity in the critical period
- PMID: 40802851
- PMCID: PMC12367176
- DOI: 10.1371/journal.pbio.3003338
Mitochondrial ROS and HIF-1α signaling mediate synaptic plasticity in the critical period
Abstract
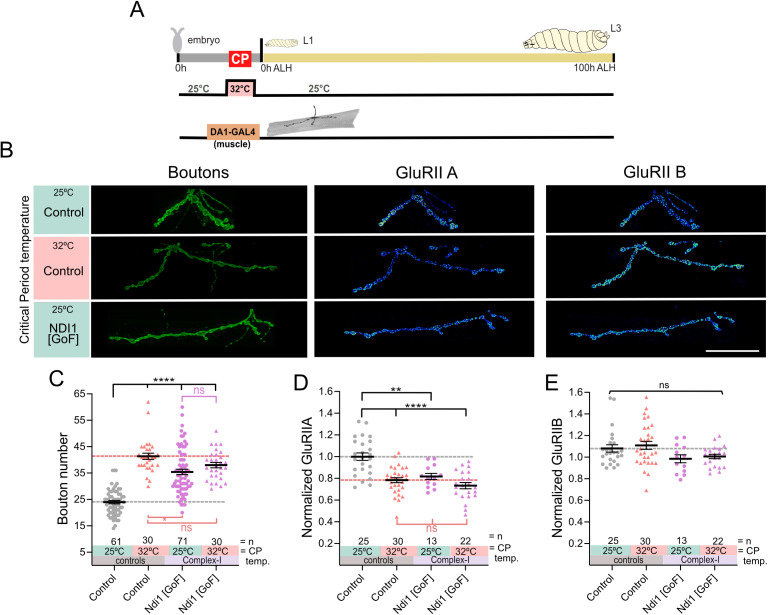
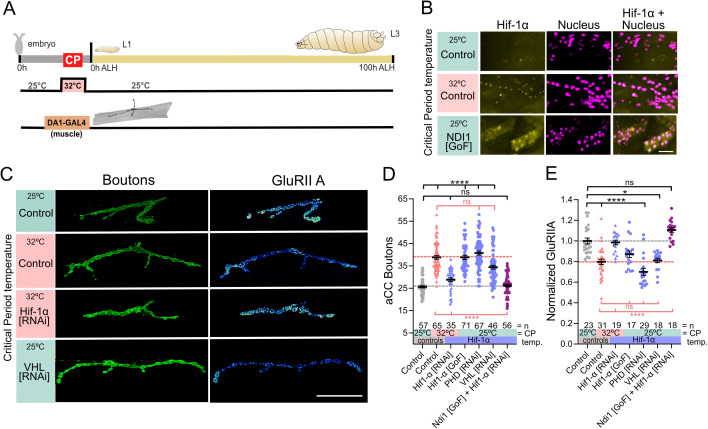
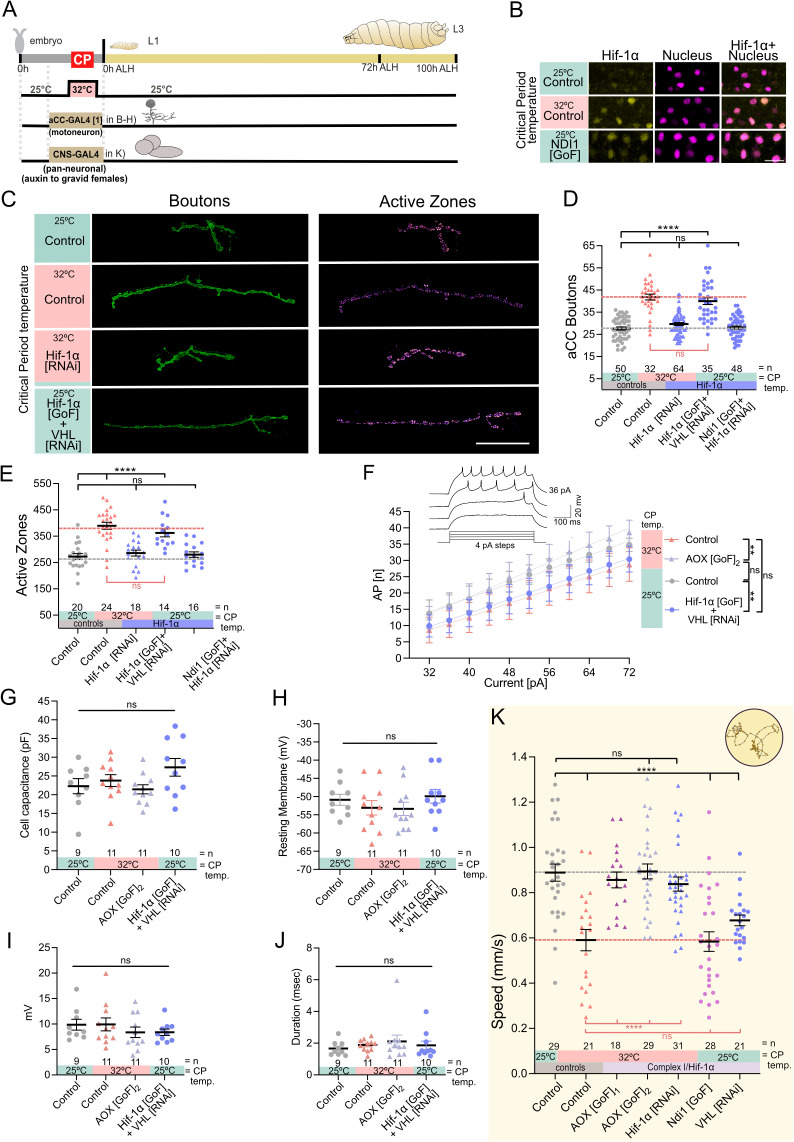
As developing networks transition from spontaneous irregular to patterned activity, they undergo plastic tuning phases, termed "critical periods"; "critical" because disturbances during these phases can lead to lasting changes in network development and output. Critical periods are common to developing nervous systems, with analogous features shared from insects to mammals, yet the core signaling mechanisms that underlie cellular critical period plasticity have remained elusive. To identify these, we exploited the Drosophila larval locomotor network as an advantageous model system. It has a defined critical period and offers unparalleled access to identified network elements, including the neuromuscular junction as a model synapse. We find that manipulations of a single motoneuron or muscle cell during the critical period lead to predictable, and permanent, cell-specific changes. This demonstrates that critical period adjustments occur at a single-cell level. Mechanistically, we identified mitochondrial reactive oxygen species (ROS) as causative. Specifically, we show that ROS produced by Complex-I of the mitochondrial electron transport chain, generated by the reverse flow of electrons, is necessary and instructive for critical period-regulated plasticity. Downstream of ROS, we identified the Drosophila homologue of hypoxia-inducible factor (HIF-1α), as required for transducing the mitochondrial ROS signal to the nucleus. This signaling axis is also sufficient to cell autonomously specify changes in neuronal properties and animal behavior but, again, only when activated during the embryonic critical period. Thus, we have identified specific mitochondrial ROS and HIF-1α as primary signals that mediate critical period plasticity.
Copyright: © 2025 Sobrido-Cameán et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



Similar articles
-
Prescription of Controlled Substances: Benefits and Risks.2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 30726003 Free Books & Documents.
-
Peripuberty Is a Sensitive Period for Prefrontal Parvalbumin Interneuron Activity to Impact Adult Cognitive Flexibility.Dev Neurosci. 2025;47(2):127-138. doi: 10.1159/000539584. Epub 2024 Jun 3. Dev Neurosci. 2025. PMID: 38830346 Free PMC article.
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
-
Maternal and neonatal outcomes of elective induction of labor.Evid Rep Technol Assess (Full Rep). 2009 Mar;(176):1-257. Evid Rep Technol Assess (Full Rep). 2009. PMID: 19408970 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
