

Distinct specificity and functions of PRC2 subcomplexes in human stem cells and cardiac differentiation
- PMID: 40803325
- PMCID: PMC12352605
- DOI: 10.1016/j.molcel.2025.07.014
Distinct specificity and functions of PRC2 subcomplexes in human stem cells and cardiac differentiation
Abstract
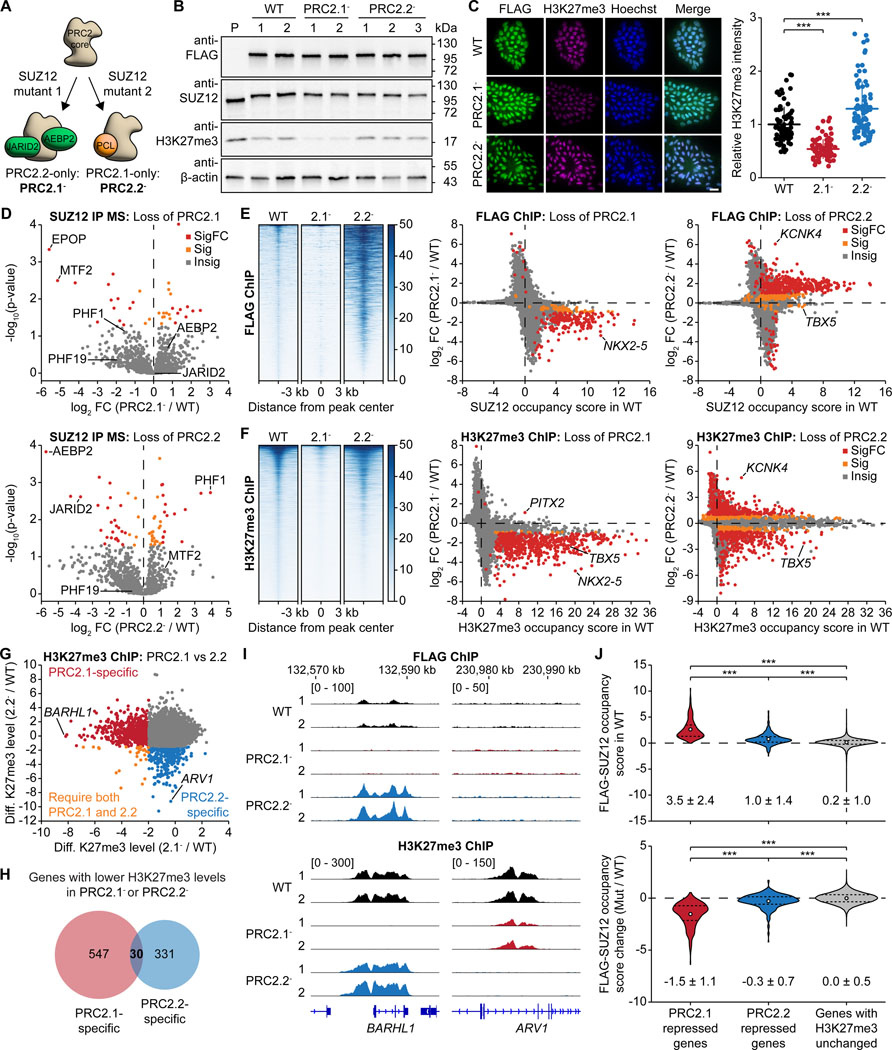
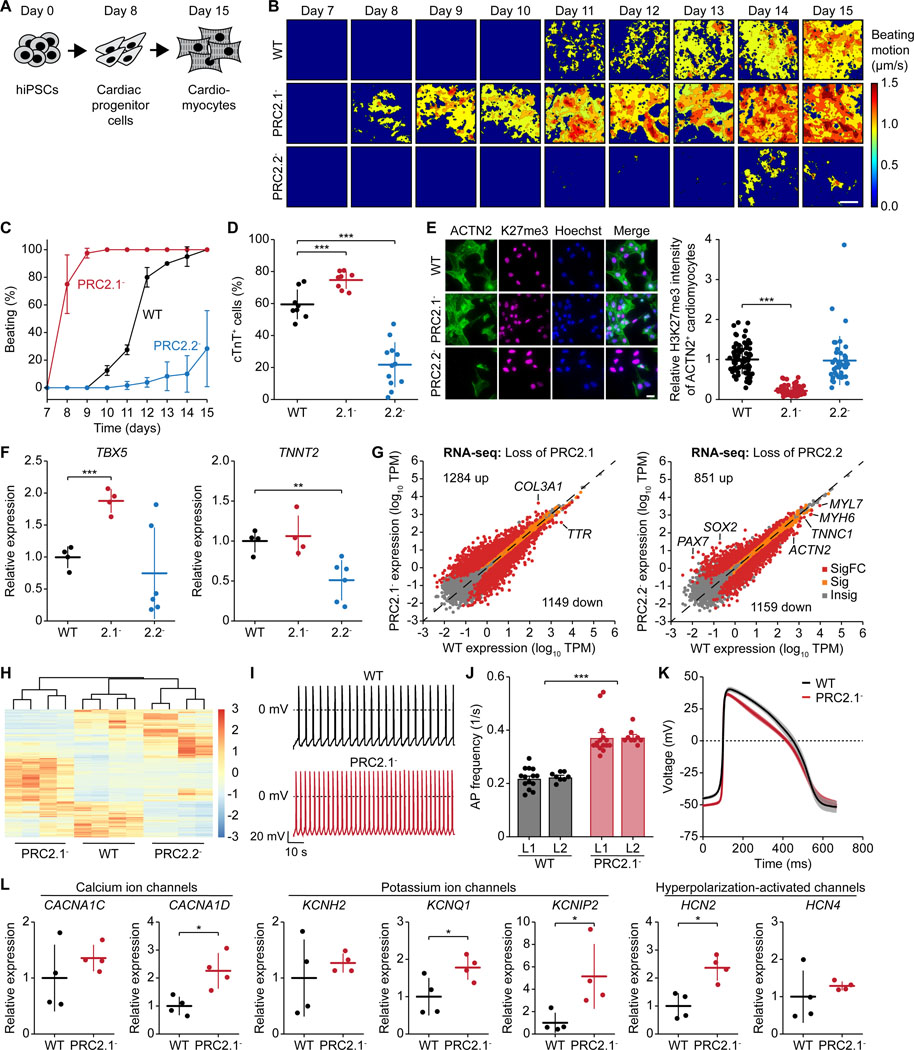
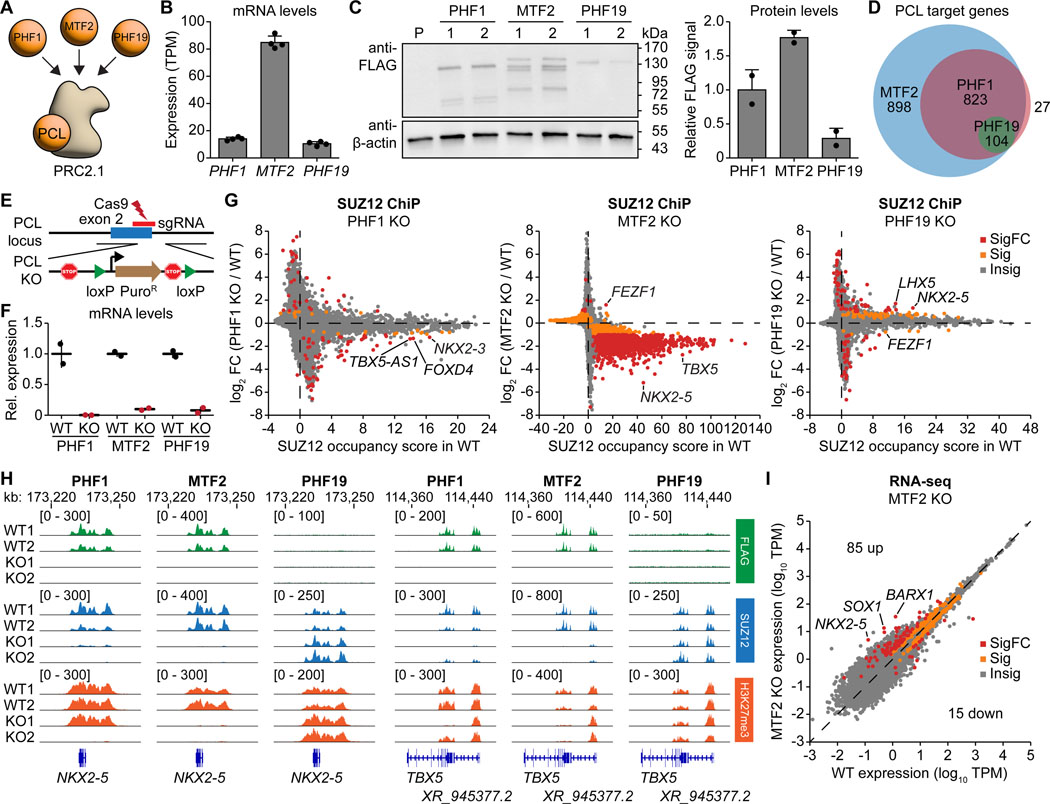
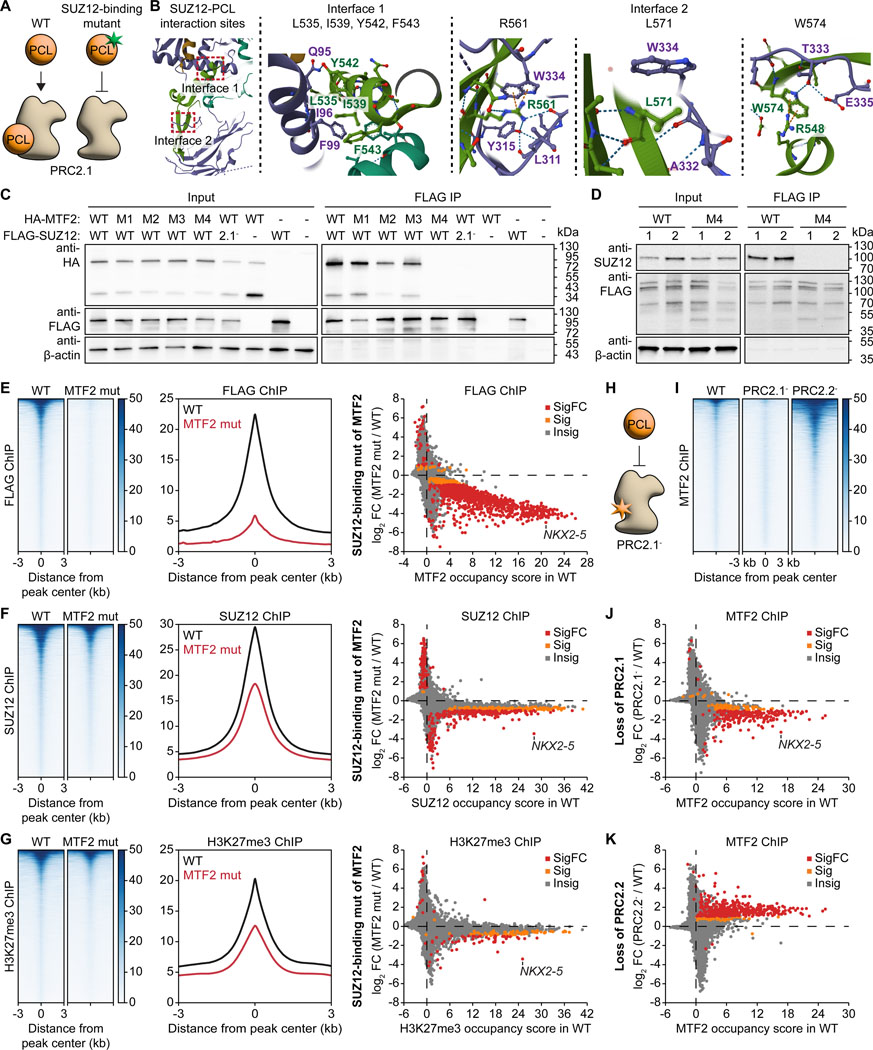
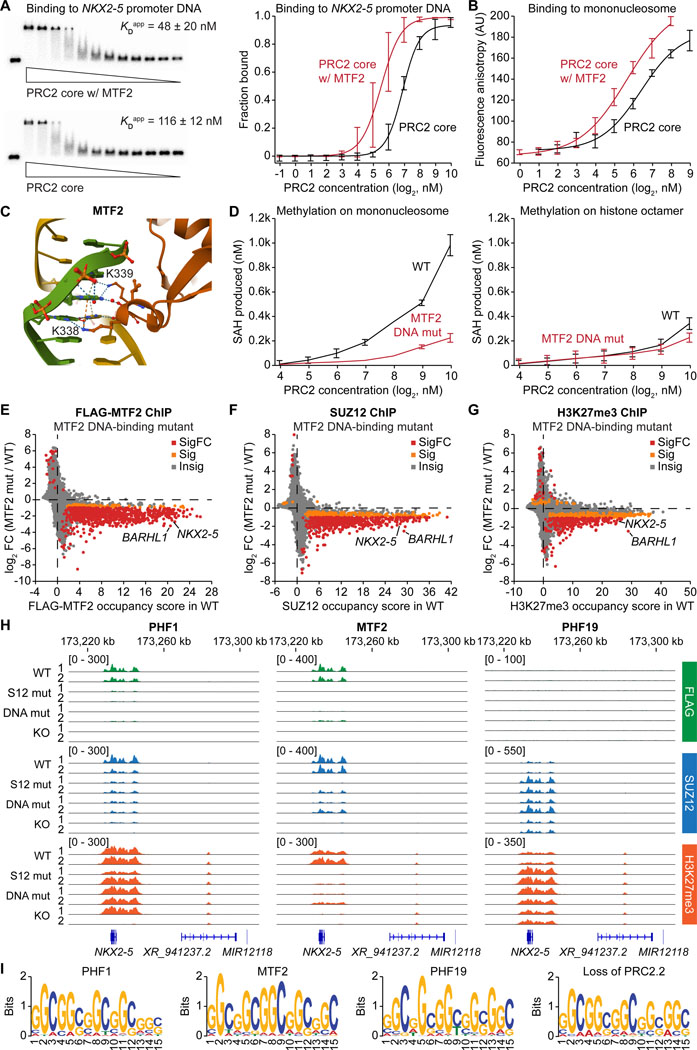
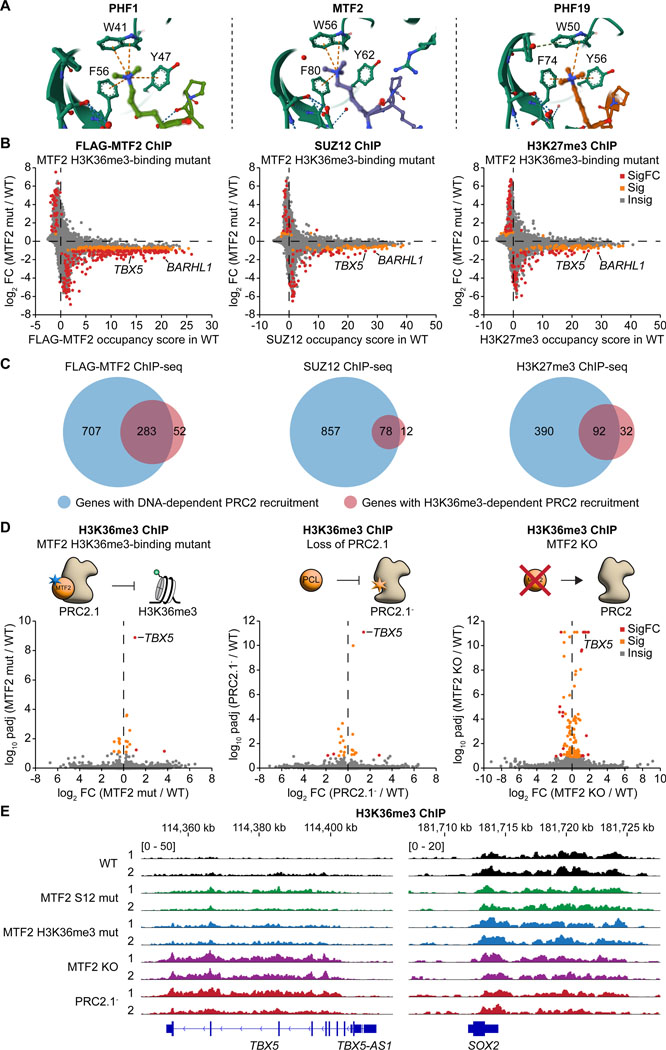
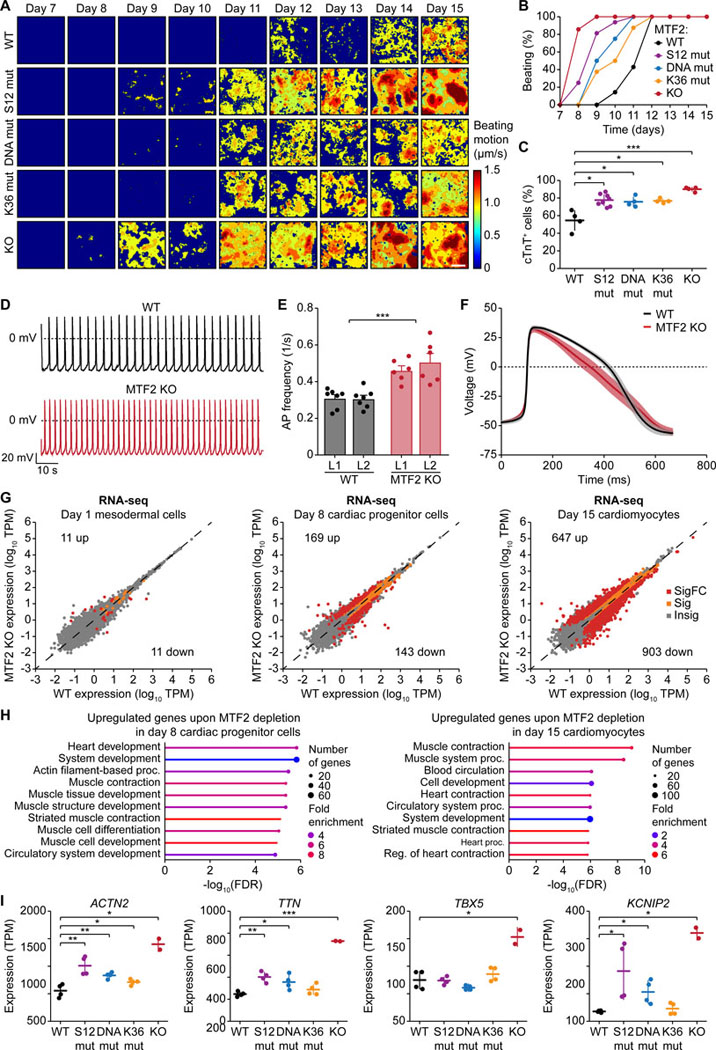
The dynamic regulation of epigenetic states relies on complex macromolecular interactions. PRC2, the methyltransferase complex depositing H3K27me3, interacts with distinct accessory proteins to form the mutually exclusive subcomplexes PHF1-PRC2.1, MTF2-PRC2.1, PHF19-PRC2.1, and PRC2.2. The functions of these subcomplexes are thought to be largely redundant. Here, we show that PRC2 subcomplexes have distinct roles in epigenetic repression of lineage-specific genes and stem cell differentiation. Using human pluripotent stem cells, we engineered a comprehensive set of separation-of-function mutants to dissect the roles of individual protein-protein and DNA-protein interactions. Our results show that PRC2.1 and PRC2.2 deposit H3K27me3 locus-specifically, resulting in opposing outcomes in cardiomyocyte differentiation. We find that MTF2 stimulates PRC2.1-mediated repression in stem cells and cardiac differentiation through its interaction with DNA and H3K36me3, while PHF19 antagonizes it. Together, these results reveal the importance and specificity of individual macromolecular interactions in Polycomb-mediated epigenetic repression in human stem cells and differentiation.
Keywords: H3K27me3; PRC2; Polycomb; cardiac differentiation; epigenetics; human pluripotent stem cells.
Copyright © 2025 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
