

Multivalent interactions with CCR4-NOT and PABPC1 determine mRNA repression efficiency by tristetraprolin
- PMID: 40804313
- PMCID: PMC12350847
- DOI: 10.1038/s41467-025-62741-7
Multivalent interactions with CCR4-NOT and PABPC1 determine mRNA repression efficiency by tristetraprolin
Abstract
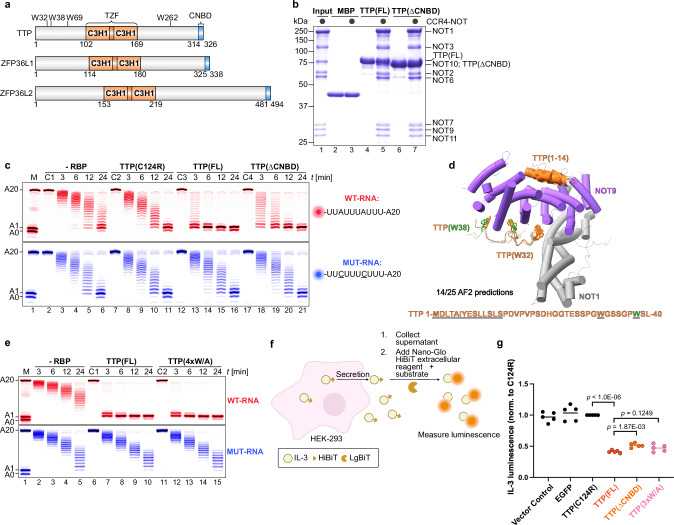
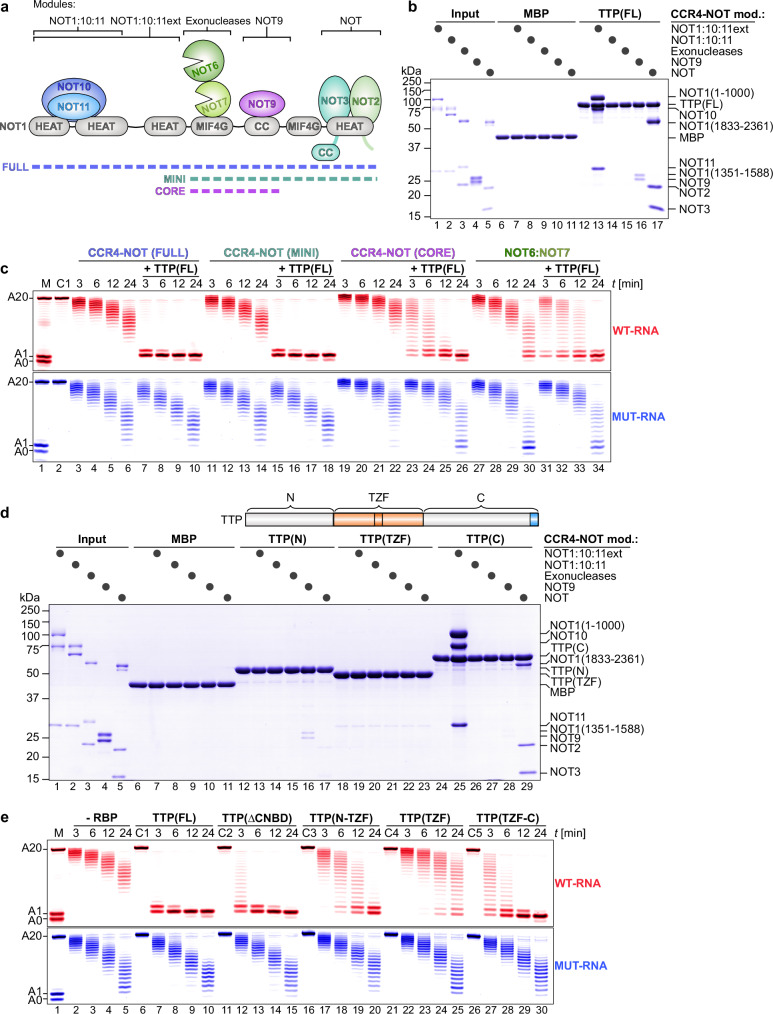
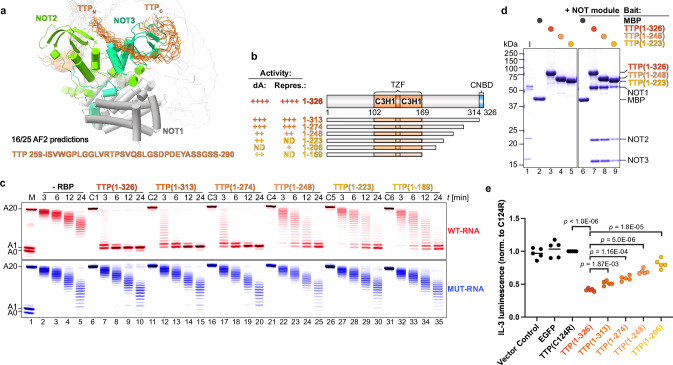
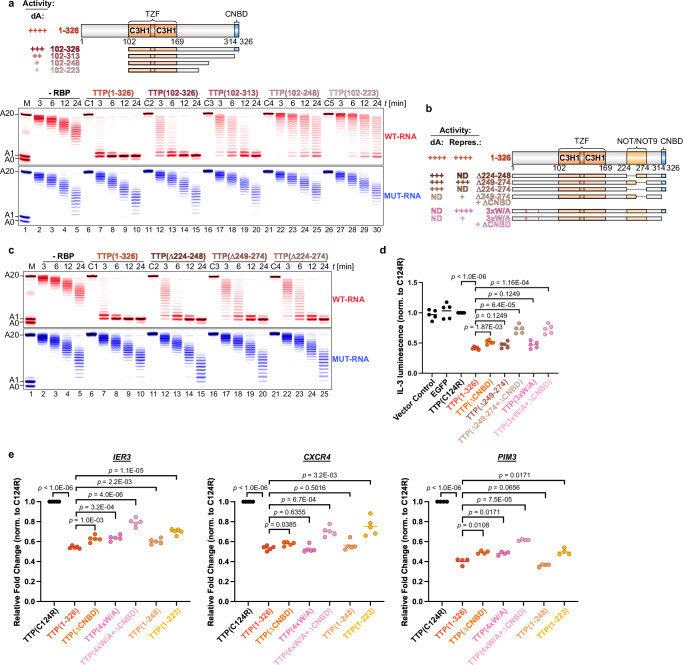
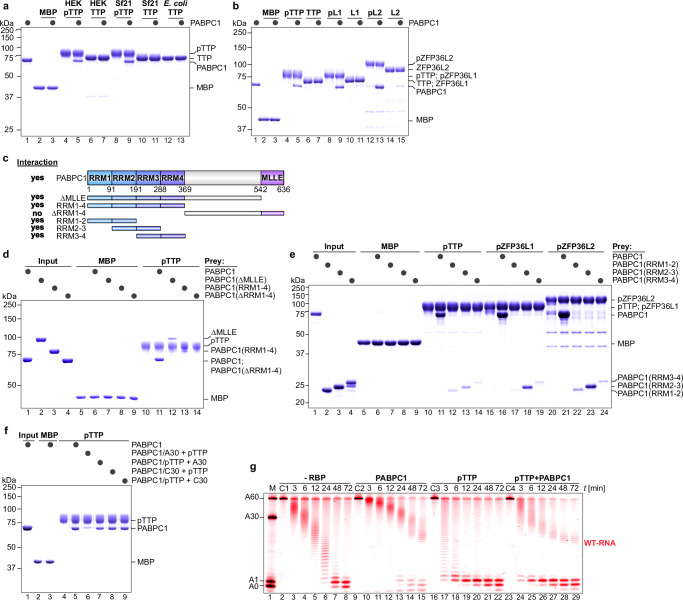
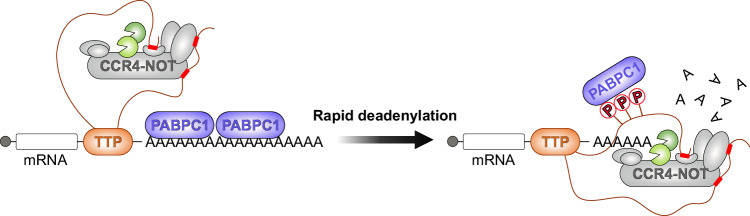
Tristetraprolin family of proteins regulate mRNA stability by binding to specific AU-rich elements in transcripts. This binding promotes the shortening of the mRNA poly(A) tail, or deadenylation, initiating mRNA degradation. The CCR4-NOT complex plays a central role in deadenylation, while the cytoplasmic poly(A)-binding protein PABPC1 typically protects mRNAs from decay. Here, we investigate how tristetraprolin interacts with CCR4-NOT and PABPC1 to control mRNA stability. Using purified proteins and in vitro assays, we find that tristetraprolin engages CCR4-NOT through multiple interaction sites and promotes its activity, emphasizing the importance of multivalent binding for efficient deadenylation. Phosphorylation of tristetraprolin does not affect its interaction with CCR4-NOT or its deadenylation activity, but is essential for tristetraprolin's binding to PABPC1. We propose that tristetraprolin promotes the processive deadenylation activity of CCR4-NOT on mRNAs containing AU-rich elements, with phosphorylation-dependent interactions with PABPC1 potentially enhancing deadenylation and promoting regulated mRNA decay.
© 2025. This is a U.S. Government work and not under copyright protection in the US; foreign copyright protection may apply.
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures

Similar articles
-
TOB Proteins Repress Translation via the CCR4-NOT Deadenylase Complex Independent of Deadenylation.Genes Cells. 2025 Sep;30(5):e70042. doi: 10.1111/gtc.70042. Genes Cells. 2025. PMID: 40755205
-
Structural basis for the recruitment of the human CCR4-NOT deadenylase complex by tristetraprolin.Nat Struct Mol Biol. 2013 Jun;20(6):735-9. doi: 10.1038/nsmb.2572. Epub 2013 May 5. Nat Struct Mol Biol. 2013. PMID: 23644599 Free PMC article.
-
Prescription of Controlled Substances: Benefits and Risks.2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 30726003 Free Books & Documents.
-
123I-MIBG scintigraphy and 18F-FDG-PET imaging for diagnosing neuroblastoma.Cochrane Database Syst Rev. 2015 Sep 29;2015(9):CD009263. doi: 10.1002/14651858.CD009263.pub2. Cochrane Database Syst Rev. 2015. PMID: 26417712 Free PMC article.
-
Signs and symptoms to determine if a patient presenting in primary care or hospital outpatient settings has COVID-19.Cochrane Database Syst Rev. 2022 May 20;5(5):CD013665. doi: 10.1002/14651858.CD013665.pub3. Cochrane Database Syst Rev. 2022. PMID: 35593186 Free PMC article.
References
LinkOut - more resources
Full Text Sources
