

Unnatural Amino Acid Photo-Crosslinking Sheds Light on Gating of the Mechanosensitive Ion Channel OSCA1.2
- PMID: 40806253
- PMCID: PMC12346119
- DOI: 10.3390/ijms26157121
Unnatural Amino Acid Photo-Crosslinking Sheds Light on Gating of the Mechanosensitive Ion Channel OSCA1.2
Abstract
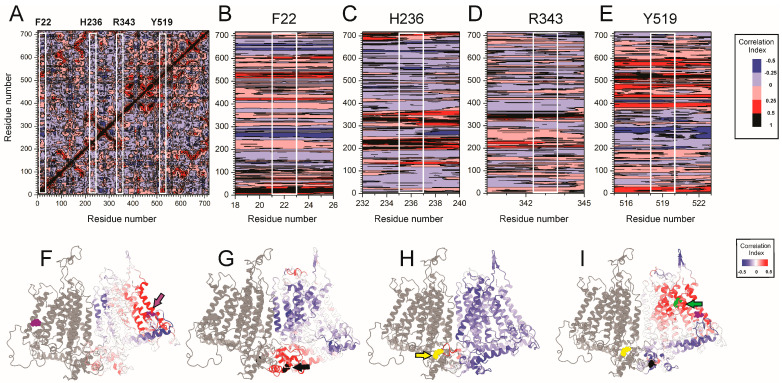
Mechanosensitive ion channels such as OSCA1.2 enable cells to sense and respond to mechanical forces by translating membrane tension into ionic flux. While lipid rearrangement in the inter-subunit cleft has been proposed as a key activation mechanism, the contributions of other domains to OSCA gating remain unresolved. Here, we combined the genetic encoding of the photoactivatable crosslinker p-benzoyl-L-phenylalanine (BzF) with functional Ca2+ imaging and molecular dynamics simulations to dissect the roles of specific residues in OSCA1.2 gating. Targeted UV-induced crosslinking at positions F22, H236, and R343 locked the channel in a non-conducting state, indicating their functional relevance. Structural analysis revealed that these residues are strategically positioned: F22 interacts with lipids near the activation gate, H236 lines the lipid-filled cavity, and R343 forms cross-subunit contacts. Together, these results support a model in which mechanical gating involves a distributed network of residues across multiple channel regions, allosterically converging on the activation gate. This study expands our understanding of mechanotransduction by revealing how distant structural elements contribute to force sensing in OSCA channels.
Keywords: OSCA1.2; ion channel gating; mechanosensing; unnatural amino acids (UAAs).
Conflict of interest statement
The authors declare no conflicts of interest.
Figures




Similar articles
-
Photoinhibition of the hERG potassium channel PAS domain by ultraviolet light speeds channel closing.Biophys J. 2024 Aug 20;123(16):2392-2405. doi: 10.1016/j.bpj.2024.05.024. Epub 2024 May 24. Biophys J. 2024. PMID: 38796698 Free PMC article.
-
Intestinal enteroendocrine cells rely on ryanodine and IP3 calcium store receptors for mechanotransduction.J Physiol. 2023 Jan;601(2):287-305. doi: 10.1113/JP283383. Epub 2022 Dec 13. J Physiol. 2023. PMID: 36428286 Free PMC article.
-
H+ and Confined Water in Gating in Many Voltage-Gated Potassium Channels: Ion/Water/Counterion/Protein Networks and Protons Added to Gate the Channel.Int J Mol Sci. 2025 Jul 29;26(15):7325. doi: 10.3390/ijms26157325. Int J Mol Sci. 2025. PMID: 40806455 Free PMC article. Review.
-
6,7-Dichloro-1H-indole-2,3-dione-3-oxime functions as a superagonist for the intermediate-conductance Ca2+-activated K+ channel KCa3.1.Mol Pharmacol. 2025 Mar;107(3):100018. doi: 10.1016/j.molpha.2025.100018. Epub 2025 Jan 31. Mol Pharmacol. 2025. PMID: 40068526
-
From gene to mechanics: a comprehensive insight into the mechanobiology of LMNA mutations in cardiomyopathy.Cell Commun Signal. 2024 Mar 27;22(1):197. doi: 10.1186/s12964-024-01546-5. Cell Commun Signal. 2024. PMID: 38539233 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
