

Orthrus: a Pumilio-family gene involved in fruiting body and dark stipe development in Coprinopsis cinerea
- PMID: 40809782
- PMCID: PMC12344735
- DOI: 10.3389/ffunb.2025.1633301
Orthrus: a Pumilio-family gene involved in fruiting body and dark stipe development in Coprinopsis cinerea
Abstract
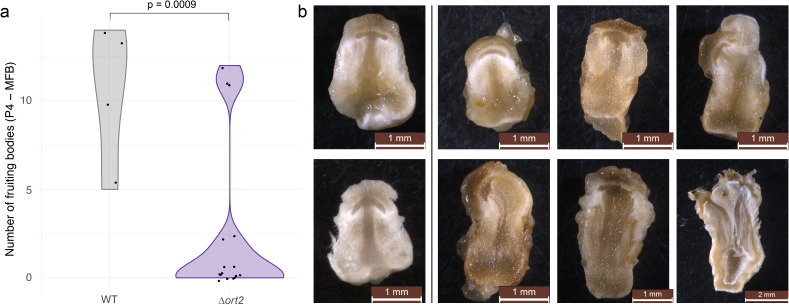
Fruiting bodies of mushroom-forming fungi (Agaricomycetes) are complex multicellular structures whose formation is regulated by a developmental program that dynamically responds to environmental changes, such as light intensity. However, the genetic architecture and regulation of this developmental program are poorly known. Here, we characterize a novel Pumilio family gene, ort2, which influences fruiting body development, particularly the formation of dark stipes, a light-dependent alternative developmental trajectory. Phylogenetic analysis of this RNA-binding protein family in fungi revealed a distinct subfamily structure, with high conservation of each subfamily within Agaricomycetes. Reverse genetics experiments in the model species Coprinopsis cinerea revealed that ort2 disruptants produced fruiting bodies, but were deficient in dark stipe formation, whereas the overexpression mutants produced significantly more dark stipes. The gene was named after Orthrus, the two-headed dog of classical mythology, based on rare but reproducible branching fruiting body phenotypes observed upon overexpression. Our findings reveal fruiting-related functions for ort2, a novel conserved RNA-binding protein, and may serve as a novel entry point for understanding the molecular basis of dark stipe development.
Keywords: Pumilio; RNA-binding proteins; dark stipe; fruiting body formation; language model.
Copyright © 2025 Szathmári, Bálint, Hegedüs, Virágh, Hou, Liu, Wu, Földi, Gagneur, Promeuschel, Csernetics and Nagy.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The reviewer MR declared a shared affiliation with the author BB to the handling editor at the time of review.
Figures



Similar articles
-
Prescription of Controlled Substances: Benefits and Risks.2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 30726003 Free Books & Documents.
-
NOVEL INVENTION OF SPORE INDUCTION IN A SISTER SPECIES TO GROUP 4 DICTYOSTELIA.Open Res Eur. 2024 Dec 5;4:239. doi: 10.12688/openreseurope.18365.2. eCollection 2024. Open Res Eur. 2024. PMID: 39564455 Free PMC article.
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
-
Systemic treatments for metastatic cutaneous melanoma.Cochrane Database Syst Rev. 2018 Feb 6;2(2):CD011123. doi: 10.1002/14651858.CD011123.pub2. Cochrane Database Syst Rev. 2018. PMID: 29405038 Free PMC article.
-
Home treatment for mental health problems: a systematic review.Health Technol Assess. 2001;5(15):1-139. doi: 10.3310/hta5150. Health Technol Assess. 2001. PMID: 11532236
References
-
- Beijen E. P. W., Maanen M. H.v., Bergh E. S.v. d., Brouns R., Marian I. M., Vries T. J.de, et al. (2025). Transcription factors fst8, ftr3 and gat1 are regulators of the immune system of the mushroom schizophyllum commune. Fungal Genet. Biol. 179, 103987. doi: 10.1016/j.fgb.2025.103987, PMID: - DOI - PubMed
-
- Buller A. H. R. (1958). Researches on fungi. Vol (VI. New York: Hafner Publishing Co; ).
-
- Chaisaena W. (2009). Light effects on fruiting body development of wildtype in comparison to light-insensitive mutant strains of the basidiomycete coprinopsis cinerea, grazing of mites (Tyrophagus putrescentiae) on the strains and production of volatile organic compounds during fruiting body development Doctoral thesis. (Göttingen, Germany: Georg-August-University Göttingen; ). doi: 10.53846/goediss-2341 - DOI
LinkOut - more resources
Full Text Sources