

The receptor-like cytoplasmic kinase AeRLCK2 mediates Nod-independent rhizobial symbiosis in Aeschynomene legumes
- PMID: 40811611
- PMCID: PMC12379891
- DOI: 10.1093/plcell/koaf201
The receptor-like cytoplasmic kinase AeRLCK2 mediates Nod-independent rhizobial symbiosis in Aeschynomene legumes
Abstract
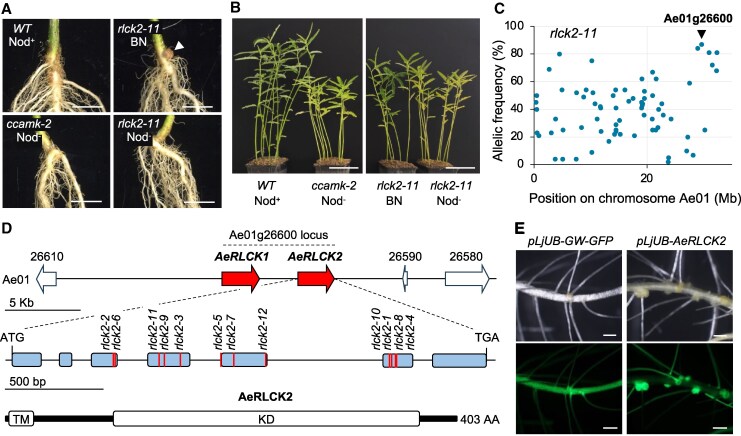
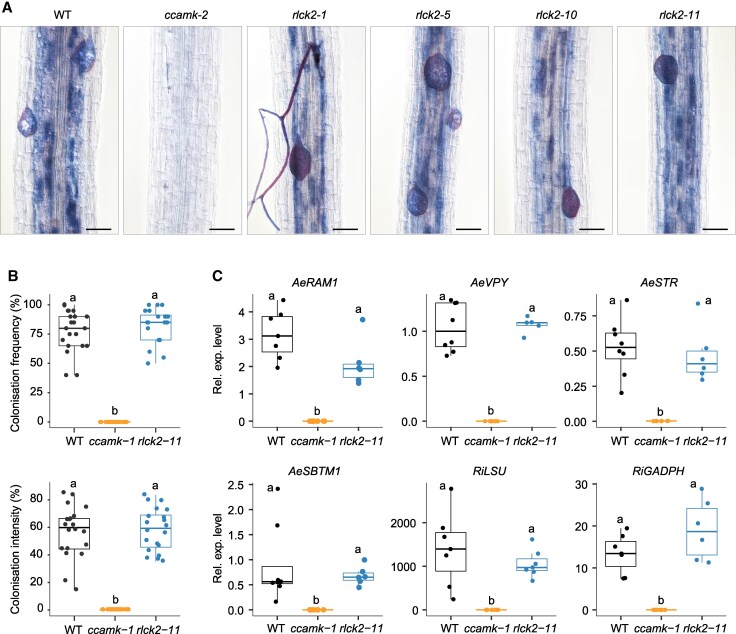
Many plants interact symbiotically with arbuscular mycorrhizal fungi to enhance inorganic phosphorus uptake, and legumes also develop a nodule symbiosis with rhizobia for nitrogen acquisition. The establishment and functioning of both symbioses rely on a common plant signaling pathway activated by structurally related Myc and Nod factors. Recently, a SPARK receptor-like kinase (RLK)/receptor-like cytoplasmic kinase (RLCK) complex was shown to be essential for arbuscular mycorrhiza formation in both monocot and dicot plants. Here, we show that in Aeschynomene legumes, the RLCK component of this receptor complex has undergone a gene duplication event and mediates a unique nodule symbiosis that is independent of rhizobial Nod factors. In Aeschynomene evenia, AeRLCK2 is crucial for nodule initiation but not for arbuscular mycorrhiza symbiosis. Additionally, AeRLCK2 physically interacts with and is phosphorylated by the cysteine-rich RLK, AeCRK, which is also required for nodulation. This finding uncovers an important molecular mechanism that controls the establishment of nodulation and is associated with Nod-independent symbiosis.
© The Author(s) 2025. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Conflict of interest statement
Conflict of interest statement. None declared.
Figures







Similar articles
-
The rhizobial type III effectors ErnA and Sup3 hijack the SUMOylation pathway to trigger nodule formation in Aeschynomene species.New Phytol. 2025 Aug;247(4):1826-1836. doi: 10.1111/nph.70334. Epub 2025 Jun 22. New Phytol. 2025. PMID: 40545786 Free PMC article.
-
A mutant-based analysis of the establishment of Nod-independent symbiosis in the legume Aeschynomene evenia.Plant Physiol. 2022 Sep 28;190(2):1400-1417. doi: 10.1093/plphys/kiac325. Plant Physiol. 2022. PMID: 35876558 Free PMC article.
-
A kinase mediator of rhizobial symbiosis and immunity in Medicago.Nature. 2025 Jul;643(8072):768-775. doi: 10.1038/s41586-025-09057-0. Epub 2025 May 6. Nature. 2025. PMID: 40328313
-
Oxygen and derived reactive species in legume-rhizobia interactions: paradoxes and dual roles.J Exp Bot. 2025 Sep 3;76(13):3758-3773. doi: 10.1093/jxb/eraf160. J Exp Bot. 2025. PMID: 40314411 Review.
-
Regulation of Root Nodule Symbiosis by Soybean Rj Genotypes and Rhizobial Effectors.Microbes Environ. 2025;40(3). doi: 10.1264/jsme2.ME25027. Microbes Environ. 2025. PMID: 40850781 Review.
References
-
- Arrighi JF, Cartieaux F, Brown SC, Rodier-Goud M, Boursot M, Fardoux J, Patrel D, Gully D, Fabre S, Chaintreuil C, et al. Aeschynomene evenia, a model plant for studying the molecular genetics of the Nod-independent rhizobium-legume symbiosis. Mol Plant Microbe Interact. 2012:25(7):851–861. 10.1094/MPMI-02-12-0045-TA - DOI - PubMed
-
- Bonaldi K, Gargani D, Prin Y, Fardoux J, Gully D, Nouwen N, Goormachtig S, Giraud E. Nodulation of Aeschynomene afraspera and A. indica by photosynthetic Bradyrhizobium sp. strain ORS285: the nod-dependent versus the nod-independent symbiotic interaction. Mol Plant Microbe Interact. 2011:24(11):1359–1371. 10.1094/MPMI-04-11-0093 - DOI - PubMed
-
- Brottier L, Chaintreuil C, Simion P, Scornavacca C, Rivallan R, Mournet P, Moulin L, Lewis GP, Fardoux J, Brown SC, et al. A phylogenetic framework of the legume genus Aeschynomene for comparative genetic analysis of the nod-dependent and nod-independent symbioses. BMC Plant Biol. 2018:18(1):333. 10.1186/s12870-018-1567-z - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
