

Elongation of the nascent avian foregut requires coordination of intrinsic and extrinsic cell behaviors
- PMID: 40812618
- PMCID: PMC12391882
- DOI: 10.1016/j.ydbio.2025.08.009
Elongation of the nascent avian foregut requires coordination of intrinsic and extrinsic cell behaviors
Abstract
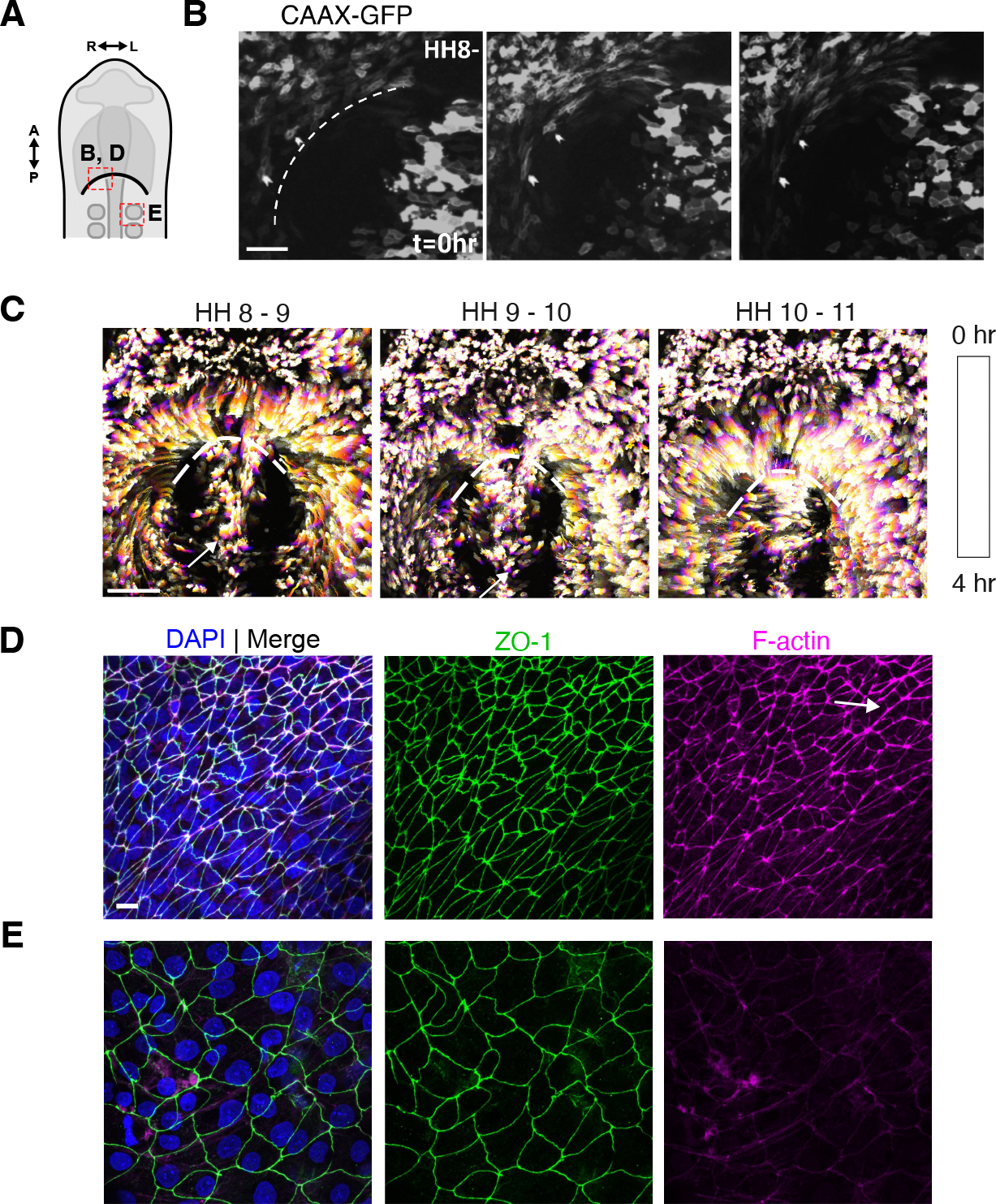
The foregut tube gives rise to the lungs and upper gastrointestinal tract, enabling vital functions of respiration and digestion. How the foregut tube forms during embryonic development has historically received considerable attention, but over the past few decades this question has primarily been addressed indirectly through studies on morphogenesis of the primitive heart tube, a closely related process. As a result, many aspects of foregut development remain unresolved. Here, we exploit the accessibility of the chick embryo to study the initial formation of the foregut tube, combining embryology with fate mapping, live imaging, and biomechanical analyses. The present study reveals that the foregut forms and elongates over a narrower time window than previously thought, and displays marked dorso-ventral and left-right asymmetries early in its development. Through tissue-specific ablation of endoderm along the anterior intestinal portal, we confirm its central role in driving foregut morphogenesis, despite not directly contributing cells to the elongating tube. We further confirm the important role of this cell population in formation of the heart tube, with evidence that this role extends to later stages of cardiac looping as well. Together, these data reveal the need for an intricate balance between intrinsic cell behaviors and extrinsic cues for normal foregut elongation, and set the stage for future studies aimed at understanding the underlying molecular cues that coordinate this balance.
Keywords: Biomechanics; Chick; Endoderm; Heart tube; Live imaging; Morphogenesis.
Copyright © 2025 Elsevier Inc. All rights reserved.
Figures





Update of
-
Elongation of the nascent avian foregut requires coordination of intrinsic and extrinsic cell behaviors.bioRxiv [Preprint]. 2024 Nov 1:2024.10.31.621372. doi: 10.1101/2024.10.31.621372. bioRxiv. 2024. Update in: Dev Biol. 2025 Nov;527:277-288. doi: 10.1016/j.ydbio.2025.08.009. PMID: 39554178 Free PMC article. Updated. Preprint.
Similar articles
-
Elongation of the nascent avian foregut requires coordination of intrinsic and extrinsic cell behaviors.bioRxiv [Preprint]. 2024 Nov 1:2024.10.31.621372. doi: 10.1101/2024.10.31.621372. bioRxiv. 2024. Update in: Dev Biol. 2025 Nov;527:277-288. doi: 10.1016/j.ydbio.2025.08.009. PMID: 39554178 Free PMC article. Updated. Preprint.
-
Prescription of Controlled Substances: Benefits and Risks.2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 30726003 Free Books & Documents.
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
-
Aspects of Genetic Diversity, Host Specificity and Public Health Significance of Single-Celled Intestinal Parasites Commonly Observed in Humans and Mostly Referred to as 'Non-Pathogenic'.APMIS. 2025 Sep;133(9):e70036. doi: 10.1111/apm.70036. APMIS. 2025. PMID: 40923351 Free PMC article. Review.
-
Signs and symptoms to determine if a patient presenting in primary care or hospital outpatient settings has COVID-19.Cochrane Database Syst Rev. 2022 May 20;5(5):CD013665. doi: 10.1002/14651858.CD013665.pub3. Cochrane Database Syst Rev. 2022. PMID: 35593186 Free PMC article.
References
-
- Adelmann Howard B. 1922. “The Significance of the Prechordal Plate: An Interpretative Study.” American Journal of Anatomy 31 (1): 55–101. 10.1002/aja.1000310104. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
