

GRSF1 loss in THP-1 macrophages promotes senescence-associated transcription in neighboring fibroblasts
- PMID: 40813599
- PMCID: PMC12354690
- DOI: 10.1038/s41598-025-11385-0
GRSF1 loss in THP-1 macrophages promotes senescence-associated transcription in neighboring fibroblasts
Abstract
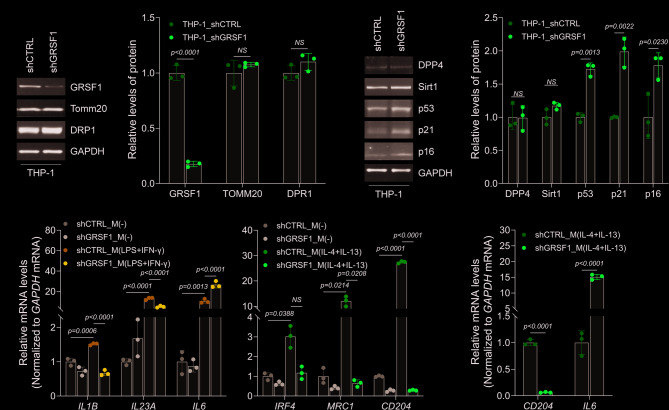
Immunosenescence, the age-associated decline in immune function, is accompanied by altered macrophage phenotypes and increased chronic inflammation. Here, we examined the role of the mitochondrial RNA-binding protein GRSF1 in regulating macrophage-driven inflammation and its impact on neighboring fibroblasts. We found that macrophages differentiated from GRSF1-deficient THP-1 monocytes, particularly M(IL-4 + IL-13) macrophages, displayed elevated IL6 mRNA expression levels and TNF-α secretion, without inducing overt senescence in macrophages themselves. Conditioned media from these macrophages triggered robust senescence-associated transcriptional changes in fibroblasts, including increased expression of IL6, TNF, DPP4, and IL8, as well as elevated SA-β-gal activity. Notably, expression of NF-κB-regulated long noncoding RNAs, such as ANRIL and PACER, was also induced in fibroblasts, suggesting the engagement of an NF-κB-linked inflammatory program. These transcriptional responses were mitigated by red ginseng extract, an anti-inflammatory compound known to suppress TNF-α signaling. Collectively, our findings suggest that GRSF1 depletion in macrophages contributes to a paracrine inflammatory niche that promotes senescence-associated gene expression in surrounding cells.
Keywords: Cell senescence; GRSF1; THP-1 macrophages.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Competing interests: The authors declare no competing interests.
Figures


Similar articles
-
Mahonia bealei (Fort.) Carr. Leaf extract modulates the TLR2/MyD88/NF-κB signaling pathway to inhibit PGN-induced inflammation in RAW264.7 cells.J Ethnopharmacol. 2025 Mar 26;344:119510. doi: 10.1016/j.jep.2025.119510. Epub 2025 Feb 17. J Ethnopharmacol. 2025. PMID: 39971016
-
SARS-CoV-2-Induced PAPOLG Contributes to Inflammation Response by Stabilizing NF-κB mRNA.J Med Virol. 2025 Jul;97(7):e70478. doi: 10.1002/jmv.70478. J Med Virol. 2025. PMID: 40607946
-
The alternative polyadenylation regulator CFIm25 promotes macrophage differentiation and activates the NF-κB pathway.Cell Commun Signal. 2025 Feb 28;23(1):115. doi: 10.1186/s12964-025-02114-1. Cell Commun Signal. 2025. PMID: 40022203 Free PMC article.
-
Unraveling the significance of innate inflammation in vascular disease.Int Rev Immunol. 2025;44(5):229-244. doi: 10.1080/08830185.2025.2489346. Epub 2025 Apr 21. Int Rev Immunol. 2025. PMID: 40255209 Review.
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2017 Dec 22;12(12):CD011535. doi: 10.1002/14651858.CD011535.pub2. Cochrane Database Syst Rev. 2017. Update in: Cochrane Database Syst Rev. 2020 Jan 9;1:CD011535. doi: 10.1002/14651858.CD011535.pub3. PMID: 29271481 Free PMC article. Updated.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
