

Functionally deficient UBOX5 variants and primary angle-closure glaucoma
- PMID: 40817263
- PMCID: PMC12356834
- DOI: 10.1038/s41467-025-62775-x
Functionally deficient UBOX5 variants and primary angle-closure glaucoma
Abstract
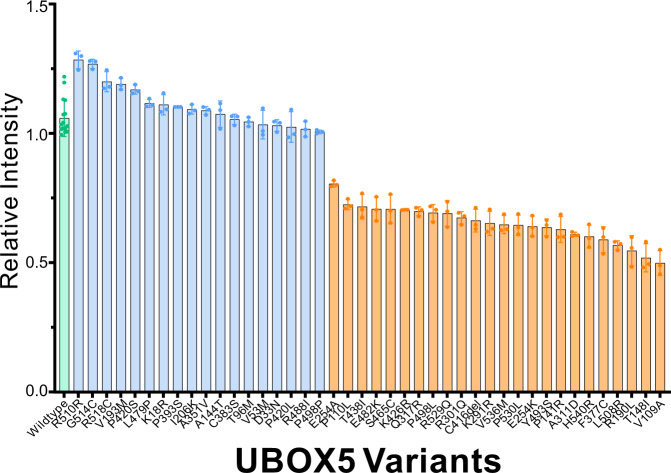
Primary angle-closure glaucoma is a major cause of irreversible blindness worldwide afflicting >20 million people. Through whole exome sequencing, we analysed the association between gene-based burden of rare, protein-altering genetic variants and disease risk in 4,667 affected individuals and 5,473 unaffected controls. We tested genes surpassing exome-wide significance (P < 2.5 × 10-6) for replication in a further 2,519 cases and 472,189 controls. We observed carriers of rare, protein-altering variants at UBOX5 (observed in 154 out of 7,186 affected individuals [2.1%] and in 3,975 out of 477,197 unaffected controls [0.83%]) to be associated with 2.13-fold increased risk of PACG (95%ci, 1.69 - 2.69; P = 1.25 × 10-10). We performed substrate trapping assays coupled with mass spectrometry and observed Binding Immunoglobulin Protein (BIP) as a key substrate for UBOX5. Biological assays showed UBOX5 acts by ubiquitinating BIP. We evaluated the functional status of 35 UBOX5 variants and observed that functionally deficient variants were enriched in affected individuals compared to controls. We validated this finding in an independent collection where 3 persons carrying functionally deficient variants were observed out of 208 cases (1.4%), whereas none were observed in 600 controls. Our findings suggest the UBOX5-BIP signalling pathway might be involved in biology of primary angle-closure glaucoma.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: A.P.K has acted as a paid consultant or lecturer to Abbvie, Aerie, Allergan, Google Health, Heidelberg Engineering, Novartis, Reichert, Santen, Thea and Topcon. The remaining authors declare no competing interests.
Figures




References
MeSH terms
LinkOut - more resources
Full Text Sources
