

Fully Synthetic Hydrogels Promote Robust Crypt Formation in Intestinal Organoids
- PMID: 40817630
- PMCID: PMC12412419
- DOI: 10.1002/adma.202509672
Fully Synthetic Hydrogels Promote Robust Crypt Formation in Intestinal Organoids
Abstract
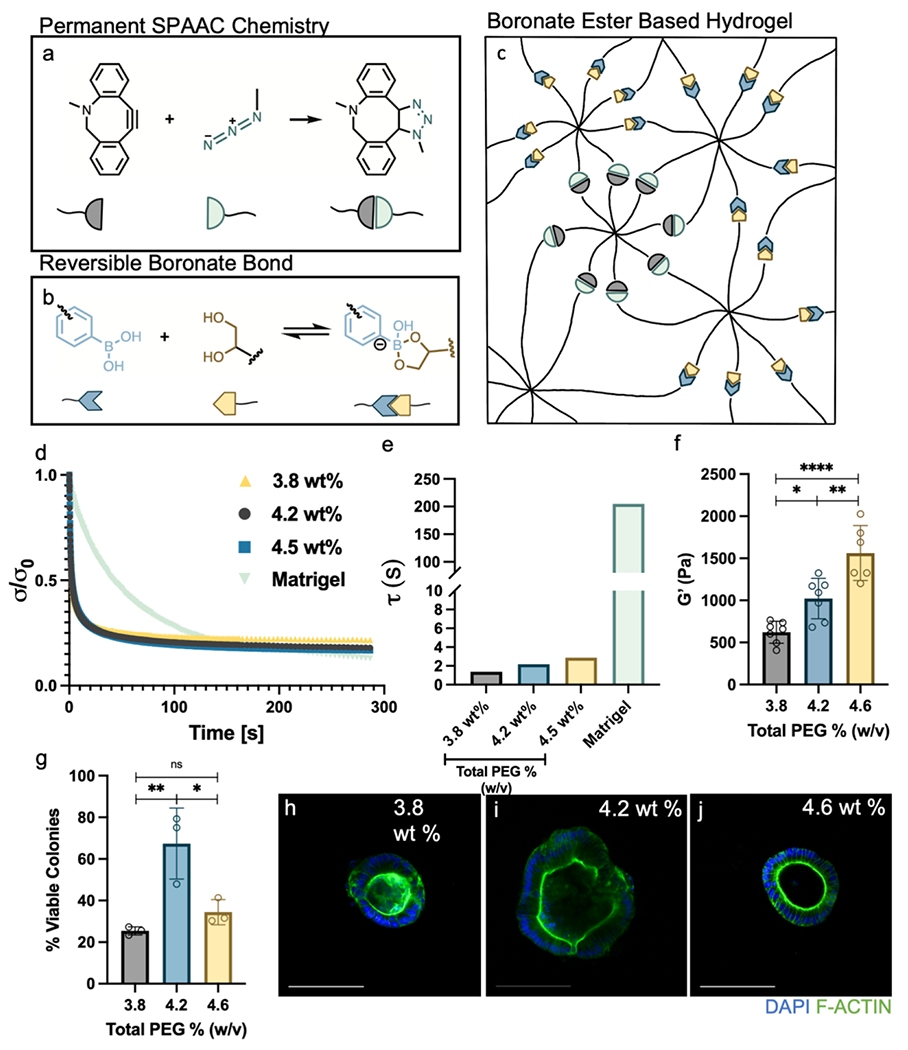
Initial landmark studies in the design of synthetic hydrogels for intestinal organoid culture identify precise matrix requirements for differentiation, namely decompression of matrix-imposed forces and supplementation of laminin. But beyond stating the necessity of laminin, organoid-laminin interactions have gone largely unstudied, as this ubiquitous requirement of exogenous laminin hinders investigation. In this work, a fast stress relaxing, boronate ester-based synthetic hydrogel is used for the culture of intestinal organoids, and it is fortuitously discovered that unlike all other synthetic hydrogels to date, laminin does not need to be supplemented for crypt formation. This highly defined material provides a unique opportunity to investigate laminin-organoid interactions and how it influences crypt evolution and organoid function. Via fluorescent labeling of non-canonical amino acids, it is further shown that adaptable boronate ester bonds increase deposition of nascent proteins, including laminin. Collectively, these results advance the understanding of how mechanical and matricellular signaling influence intestinal organoid development.
Keywords: biomaterials; extracellular matrix; intestinal organoids; laminin; stress relaxing hydrogels.
© 2025 Wiley‐VCH GmbH.
Conflict of interest statement
Conflicts of Interest Disclosure
The authors have no conflicts to report.
Figures





Update of
-
Fully synthetic hydrogels promote robust crypt formation in intestinal organoids.bioRxiv [Preprint]. 2024 Jul 10:2024.07.06.602364. doi: 10.1101/2024.07.06.602364. bioRxiv. 2024. Update in: Adv Mater. 2025 Oct;37(43):e09672. doi: 10.1002/adma.202509672. PMID: 39026840 Free PMC article. Updated. Preprint.
References
-
- Sato T, Vries RG, Snippert HJ, van de Wetering M, Barker N, Stange DE, van Es JH, Abo A, Kujala P, Peters PJ, Clevers H, Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche, Nature 459(7244) (2009) 262–5. - PubMed
-
- Sato T, Clevers H, Growing self-organizing mini-guts from a single intestinal stem cell: mechanism and applications, Science 340(6137) (2013) 1190–4. - PubMed
-
- Gjorevski N, Sachs N, Manfrin A, Giger S, Bragina ME, Ordonez-Moran P, Clevers H, Lutolf MP, Designer matrices for intestinal stem cell and organoid culture, Nature 539(7630) (2016) 560–564. - PubMed
MeSH terms
Substances
Grants and funding
- R01DK120921 (K.S.A.)/DK/NIDDK NIH HHS/United States
- P30-CA046934 (P.J.D.)/DK/NIDDK NIH HHS/United States
- W911NF-19-2-004/Defense Sciences Office, DARPA
- F31 DK126427/DK/NIDDK NIH HHS/United States
- P30 CA046934/CA/NCI NIH HHS/United States
- P30-DK116073 (P.J.D.)/DK/NIDDK NIH HHS/United States
- R01 DK120921/DK/NIDDK NIH HHS/United States
- K99 DK135907/DK/NIDDK NIH HHS/United States
- DARPA grant
- K99 DK135907-01 (M.R.B.)/DK/NIDDK NIH HHS/United States
- P30 DK116073/DK/NIDDK NIH HHS/United States
- DGE 2040434/National Science Foundation Graduate Research Fellowship Program
- 2033723/National Science Foundation RECODE
- F31 DK126427/DK/NIDDK NIH HHS/United States
- R01DK120921 (K.S.A.)/DK/NIDDK NIH HHS/United States
- P30-DK116073 (P.J.D.)/DK/NIDDK NIH HHS/United States
- P30-CA046934 (P.J.D.)/DK/NIDDK NIH HHS/United States
- K99 DK135907-01 (M.R.B.)/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
