

Pangenome analysis of transposable element insertion polymorphisms reveals features underlying cold tolerance in rice
- PMID: 40819063
- PMCID: PMC12357961
- DOI: 10.1038/s41467-025-62887-4
Pangenome analysis of transposable element insertion polymorphisms reveals features underlying cold tolerance in rice
Abstract
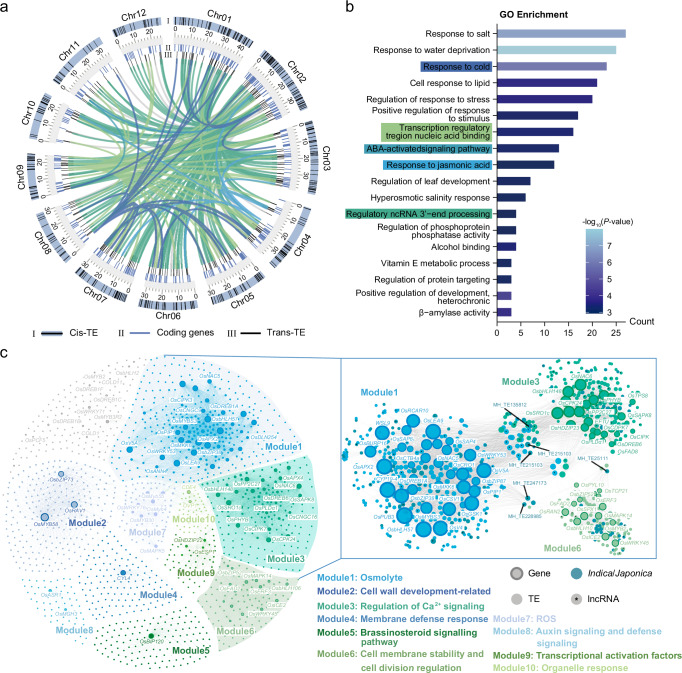
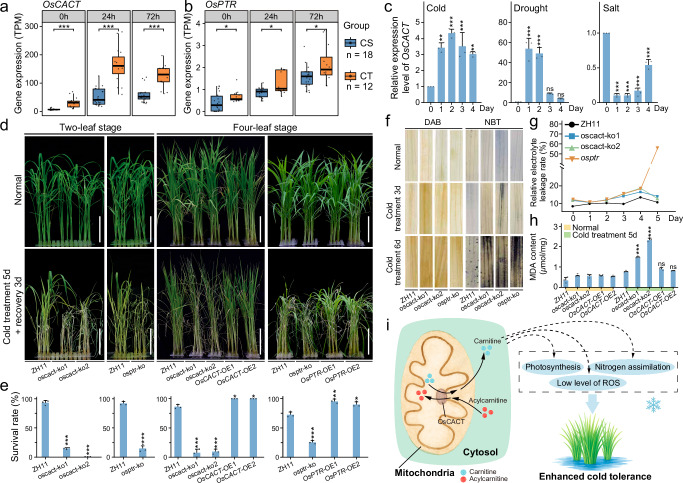
Transposable elements (TEs) introduce genetic and epigenetic variability, contributing to gene expression patterns that drive adaptive evolution in plants. Here, we investigate TE architecture and its effect on cold tolerance in rice. By analyzing a pangenome graph and the resequencing data of 165 rice accessions, we identify 30,316 transposable element insertion polymorphism (TIP) sites, highlighting significant diversity among polymorphic TEs (pTEs). We observe that pTEs exhibit increased H3K27me3 enrichment, suggesting a potential role in epigenetic differentiation under cold stress and in the transcriptional regulation of the cold response. We identify 26,914 TEs responsive to cold stress from transcriptome data, indicating their potential significance in regulatory networks for this response. Our TIP-GWAS analysis reveal two cold tolerance genes OsCACT and OsPTR. The biological functions of these genes are confirmed using knockout and overexpression lines. Our web tool ( https://cbi.gxu.edu.cn/RICEPTEDB/ ) makes all pTEs available to researchers for further analysis. These findings provide valuable targets for breeding cold-tolerant rice varieties, indicating the potential importance of pTEs in crop enhancement.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures



References
-
- Wing, R. A., Purugganan, M. D. & Zhang, Q. The rice genome revolution: from an ancient grain to Green Super Rice. Nat. Rev. Genet.19, 505–517 (2018). - PubMed
-
- Lesk, C., Rowhani, P. & Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature529, 84–87 (2016). - PubMed
-
- Fu, J. et al. Extreme rainfall reduces one-twelfth of China’s rice yield over the last two decades. Nat. Food4, 416–426 (2023). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
