

Encephalomyocarditis virus protein 2B* interacts with 14-3-3 proteins through a phosphorylated C-terminal binding motif
- PMID: 40824083
- PMCID: PMC12421828
- DOI: 10.1128/mbio.01008-25
Encephalomyocarditis virus protein 2B* interacts with 14-3-3 proteins through a phosphorylated C-terminal binding motif
Abstract
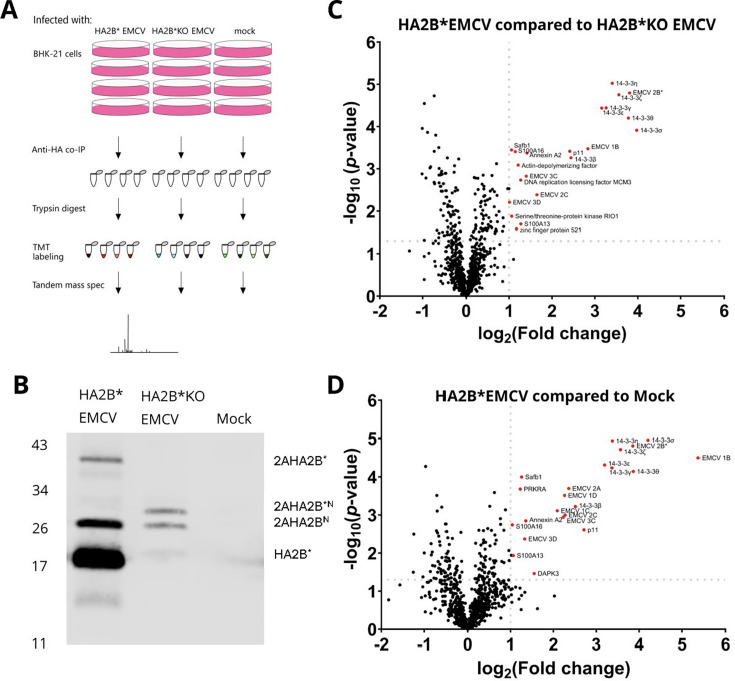
Encephalomyocarditis virus (EMCV) has been an important model RNA virus for decades. Although most of the EMCV proteins are obtained via proteolytic cleavage of a long polyprotein, 2B* is expressed from a short overlapping open reading frame via an unusual protein-stimulated temporally dependent ribosomal frameshifting mechanism. Mutant viruses that are unable to express 2B* have a small plaque phenotype due to delayed onset and inefficient progression of multiple lytic cell death pathways. However, the mechanism by which 2B* promotes these pathways has not yet been characterized. By tagging 2B* within the viral genome, we identified putative interaction partners of 2B* and showed that 2B* binds all seven members of the 14-3-3 protein family during virus infection. Binding is entirely dependent on a mode III motif containing a phosphoserine (RRNpSS) at the 2B* C terminus. This may impede other functions of the 14-3-3 proteins, including their role in promoting antiviral signaling. Ablating the 2B*:14-3-3 interaction had no effect upon plaque size, indicating that the function of this interaction is unrelated to the role of 2B* in promoting lytic cell death. This work expands our knowledge of the protein complement of this important model virus and the binding repertoire and specificity of host 14-3-3 proteins.IMPORTANCEEncephalomyocarditis virus (EMCV) infects a range of species, causing economically important reproductive disorders in pigs and encephalitis and myocarditis in rodents. Due to its wide host range, it is an important model pathogen for investigating virus-host interactions. EMCV expresses an accessory protein, 2B*, from an overlapping open reading frame via an unusual ribosomal frameshifting mechanism. Although the frameshifting mechanism has been established, the function of the 2B* protein had not been explored until recently. Here, we determined the host proteins to which 2B* binds and found that it specifically binds to all members of the 14-3-3 protein family, which, among other roles, contribute to the innate immune response to viral infection in mammalian cells. The interaction requires a specific stretch of amino acids at the end of 2B*. Binding to 2B* may reduce the opportunities for these 14-3-3 proteins to bind to host proteins and perform their usual roles; therefore, by interacting with the 14-3-3 proteins, 2B* may affect multiple host cell functions, including immune response activation.
Keywords: 14-3-3 proteins; 2B*; EMCV; cardiovirus; innate immunity; interferon beta; picornavirus; proteomics; ribosome frameshifting.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
-
- Loughran G, Libbey JE, Uddowla S, Scallan MF, Ryan MD, Fujinami RS, Rieder E, Atkins JF. 2013. Theiler’s murine encephalomyelitis virus contrasts with encephalomyocarditis and foot-and-mouth disease viruses in its functional utilization of the StopGo non-standard translation mechanism. J Gen Virol 94:348–353. doi: 10.1099/vir.0.047571-0 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
