

Structural and genetic basis of HIV-1 envelope V2 apex recognition by rhesus broadly neutralizing antibodies
- PMID: 40824240
- PMCID: PMC12379892
- DOI: 10.1084/jem.20250638
Structural and genetic basis of HIV-1 envelope V2 apex recognition by rhesus broadly neutralizing antibodies
Abstract
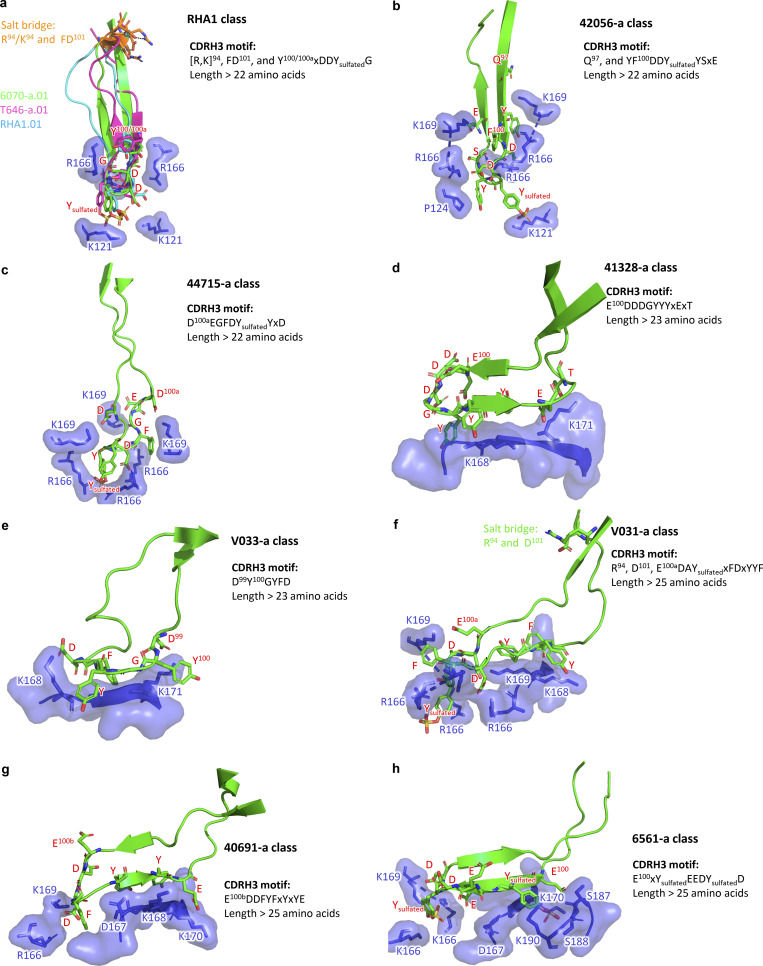
Broadly neutralizing antibodies targeting the V2 apex of HIV-1 envelope are desired as vaccine design templates, but few have been described. Here, we report 11 lineages of V2 apex-neutralizing antibodies from simian-human immunodeficiency virus (SHIV)-infected rhesus macaques and determine cryo-EM structures for 9. A single V2 apex-neutralizing lineage accounted for cross-clade breadth in most macaques, and somatic hypermutation relative to breadth was generally low, exemplified by antibody V033-a.01 with <5% nucleotide mutation and 37% breadth (208-strain panel). Envelope complex structures revealed eight different antibody classes (one multi-donor) and the complete repertoire of all five possible recognition topologies, recapitulating canonical human modes of apex insertion and C-strand hydrogen bonding. Despite this diversity in recognition, all rhesus-V2 apex antibodies were derived from reading frame two of the DH3-15*01 gene. Collectively, these results define-in rhesus-the structural and genetic basis of HIV-1 V2 apex recognition and demonstrate unprecedented structural plasticity of a highly selected immunogenetic element.
© 2025 Roark et al.
Conflict of interest statement
Disclosures: B.F. Haynes reported grants from National Institutes of Health and Bill and Melinda Gates Foundation outside the submitted work. No other disclosures were reported.
Figures













References
-
- Andrabi, R., Voss J.E., Liang C.H., Briney B., Mccoy L.E., Wu C.Y., Wong C.H., Poignard P., and Burton D.R.. 2015. Identification of common features in prototype broadly neutralizing antibodies to HIV envelope V2 apex to facilitate vaccine design. Immunity. 43:959–973. 10.1016/j.immuni.2015.10.014 - DOI - PMC - PubMed
-
- Barbian, H.J., Decker J.M., Bibollet-Ruche F., Galimidi R.P., West A.P. Jr., Learn G.H., Parrish N.F., Iyer S.S., Li Y., Pace C.S., et al. 2015. Neutralization properties of simian immunodeficiency viruses infecting chimpanzees and gorillas. mBio. 6:e00296-15. 10.1128/mBio.00296-15 - DOI - PMC - PubMed
-
- Bauer, A., Lindemuth E., Marino F.E., Krause R., Joy J., Docken S.S., Mallick S., Mccormick K., Holt C., Georgiev I., et al. 2023. Adaptation of a transmitted/founder simian-human immunodeficiency virus for enhanced replication in rhesus macaques. PLoS Pathog. 19:e1011059. 10.1371/journal.ppat.1011059 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
- INV-041767/GATES/Gates Foundation/United States
- UM1 AI 144371/Duke Consortium for HIV Vaccine Development
- R01 AI167716/AI/NIAID NIH HHS/United States
- R61 AI161818/AI/NIAID NIH HHS/United States
- R61 AI176583/AI/NIAID NIH HHS/United States
- R37 AI150590/AI/NIAID NIH HHS/United States
- INV-007939/GATES/Gates Foundation/United States
- R61 AI 176583/NH/NIH HHS/United States
- R01 AI165080/NH/NIH HHS/United States
- Duke Human Vaccine Institute
- INV-007939/GATES/Gates Foundation/United States
- UM1 AI144371/AI/NIAID NIH HHS/United States
- T32 AI007632/AI/NIAID NIH HHS/United States
- R37 AI 150590/NH/NIH HHS/United States
- National Institute of Allergy and Infectious Diseases
- R01 AI160607/NH/NIH HHS/United States
- T32-AI007632/Training Grant in HIV Pathogenesis
- P30 AI045008/AI/NIAID NIH HHS/United States
- R01 AI 050529/NH/NIH HHS/United States
- Vaccine Research Center
- R01 AI160607/AI/NIAID NIH HHS/United States
- R01 AI165080/AI/NIAID NIH HHS/United States
- R01 AI050529/AI/NIAID NIH HHS/United States
- INV-041767/GATES/Gates Foundation/United States
- R01 AI 167716/NH/NIH HHS/United States
- HI/NHLBI NIH HHS/United States
- R61 AI 161818/NH/NIH HHS/United States
LinkOut - more resources
Full Text Sources
Miscellaneous
