

Loss of YTHDF2 enhances Th9 programming and CAR-Th9 cell antitumor efficacy
- PMID: 40826275
- PMCID: PMC12614224
- DOI: 10.1038/s41590-025-02235-2
Loss of YTHDF2 enhances Th9 programming and CAR-Th9 cell antitumor efficacy
Abstract
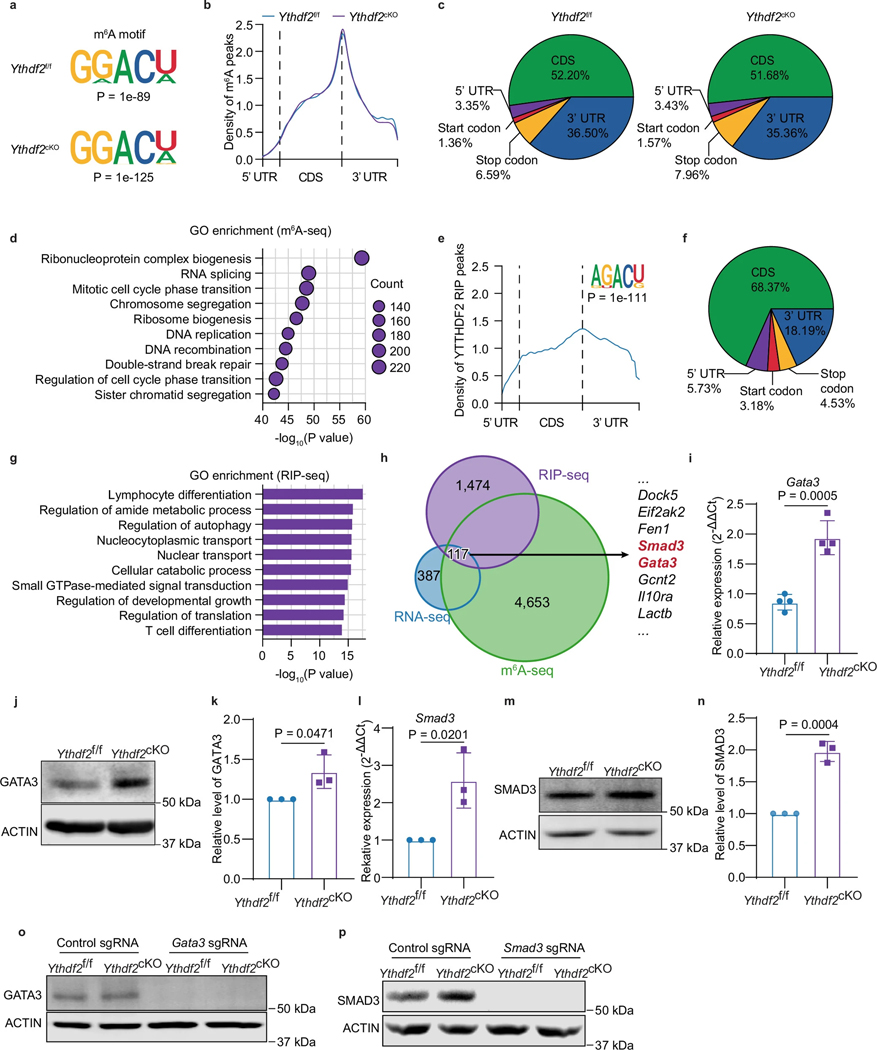
CD4+ T cells differentiate into various subsets, including T helper 1 (Th1), Th2, Th9, Th17 and regulatory T (Treg) cells, which are essential for immune responses and cancer immunotherapy. However, the role of RNA N6-methyladenosine (m6A) modification in this differentiation is unclear. Here we show that YTHDF2, an important m6A reader protein known to destabilize m6A-modified mRNA, negatively regulates Th9 cell differentiation. Ablation of Ythdf2 in both mouse and human naive CD4+ T cells promotes Th9 differentiation by stabilizing Gata3 and Smad3 mRNA under interleukin-4 (IL-4) and transforming growth factor β (TGF-β) signaling, respectively. Ythdf2-deficient Th9 cells produce increased amounts of IL-9 and IL-21, leading to increased tumor infiltration and cytotoxicity by CD8+ T cells and natural killer (NK) cells, thereby improving antitumor activity compared with wild-type Th9 cells. Moreover, YTHDF2 depletion in CAR-Th9 cells enhances their immune activation, reduces their terminal differentiation and augments their antitumor efficacy. Targeting YTHDF2 is thereby a promising strategy to enhance Th9 and CAR-Th9 cell-based cancer immunotherapies.
© 2025. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures

















References
Methods-only references
MeSH terms
Substances
Grants and funding
- R01 AI129582/AI/NIAID NIH HHS/United States
- TRAN1-14003, DISC2-14190/California Institute for Regenerative Medicine (CIRM)
- TRAN1-14716/California Institute for Regenerative Medicine (CIRM)
- R21 CA223400/CA/NCI NIH HHS/United States
- R01 CA266457/CA/NCI NIH HHS/United States
- NS106170, AI129582, CA247550, CA264512, CA266457, CA223400/Foundation for the National Institutes of Health (Foundation for the National Institutes of Health, Inc.)
- R35 CA210087/CA/NCI NIH HHS/United States
- U19 CA264512/CA/NCI NIH HHS/United States
- P30 CA033572/CA/NCI NIH HHS/United States
- P01 CA163205/CA/NCI NIH HHS/United States
- R01 CA247550/CA/NCI NIH HHS/United States
- R01 CA265095/CA/NCI NIH HHS/United States
- R01 NS106170/NS/NINDS NIH HHS/United States
- CA210087, CA265095, CA163205/Foundation for the National Institutes of Health (Foundation for the National Institutes of Health, Inc.)
LinkOut - more resources
Full Text Sources
Medical
Research Materials
