

This is a preprint.
Mapping of in vivo cleavage sites uncovers a major role for yeast RNase III in regulating protein-coding genes
- PMID: 40832349
- PMCID: PMC12363893
- DOI: 10.1101/2025.03.07.642061
Mapping of in vivo cleavage sites uncovers a major role for yeast RNase III in regulating protein-coding genes
Abstract
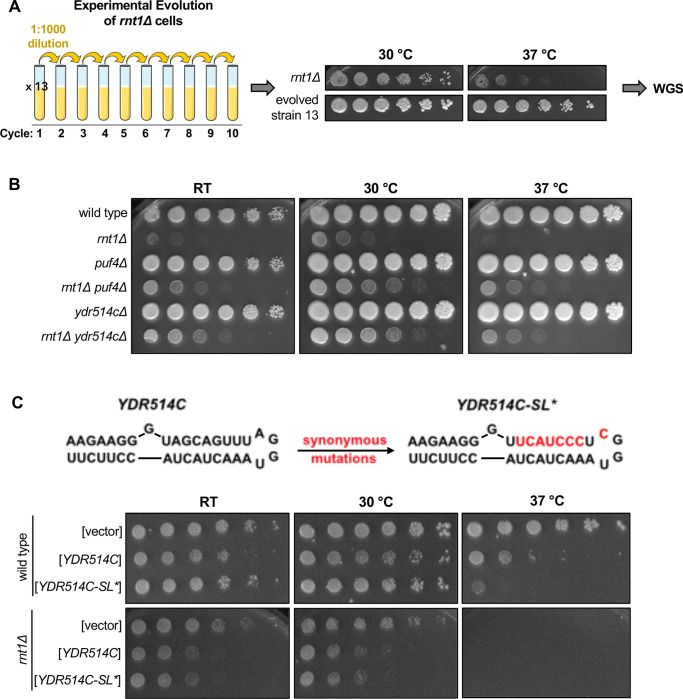
A large fraction of newly transcribed RNA is degraded in the nucleus, but nuclear mRNA degradation pathways remain largely understudied. The yeast nuclear endoribonuclease Rnt1 has a well-characterized role in the maturation of many ncRNA precursors. However, the scope and consequence of its function in mRNA degradation pathways is much less defined. Here, we take a whole-transcriptome approach to identify Rnt1 cleavage sites throughout the yeast transcriptome in vivo, at single-nucleotide resolution. We discover previously unknown Rnt1 cleavage sites in many protein-coding regions and find that the sequences and structures necessary for cleavage mirror those required for the cleavage of known targets. We show that the nuclear localization of Rnt1 functions as an additional layer of target selection control and that cleaved mRNAs are likely exported to the cytoplasm to be degraded by Xrn1. Further, we find that several cleavage products are much more abundant in our degradome sequencing libraries than decapping products, and strikingly, mutations in one Rnt1 target, YDR514C, suppress the growth defect of a RNT1 deletion. Overexpression of YDR514C results in slow growth, further suggesting that Rnt1 may limit the expression of YDR514C to maintain proper cell growth. This study uncovers a broader target range and function for the well-known RNase III enzyme.
Figures






References
-
- Abramson J., Adler J., Dunger J., Evans R., Green T., Pritzel A., Ronneberger O., Willmore L., Ballard A. J., Bambrick J., Bodenstein S. W., Evans D. A., Hung C.-C., O’Neill M., Reiman D., Tunyasuvunakool K., Wu Z., Žemgulytė A., Arvaniti E., … Jumper J. M. (2024). Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature, 630(8016), 493–500. 10.1038/s41586-024-07487-w - DOI - PMC - PubMed
-
- Andrews S. (2010). FastQC: A quality control tool for high throughput sequence data. Http://Www.Bioinformatics.Babraham.Ac.Uk/Projects/Fastqc.
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources