

SLC45A4 is a pain gene encoding a neuronal polyamine transporter
- PMID: 40836097
- PMCID: PMC12507699
- DOI: 10.1038/s41586-025-09326-y
SLC45A4 is a pain gene encoding a neuronal polyamine transporter
Abstract
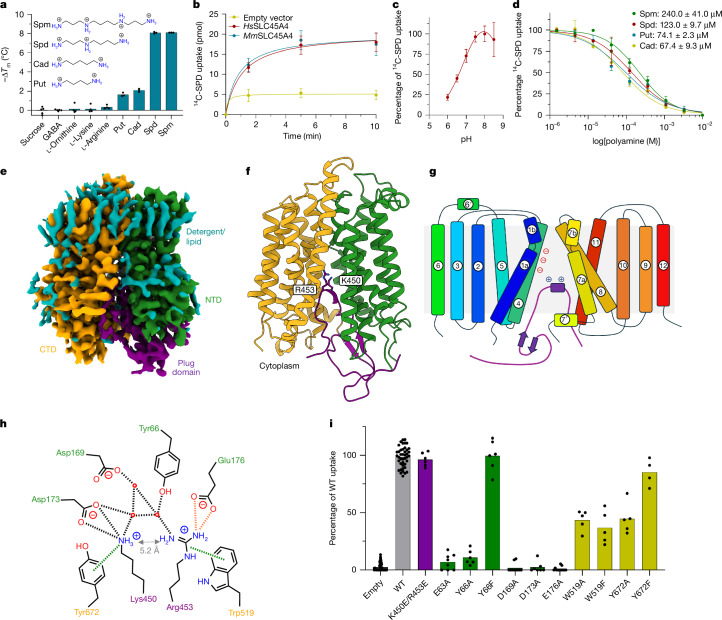
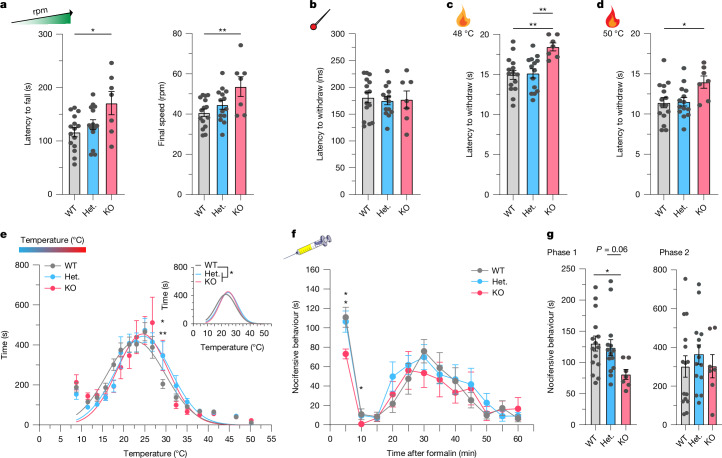
Polyamines are regulatory metabolites with key roles in transcription, translation, cell signalling and autophagy1. They are implicated in multiple neurological disorders, including stroke, epilepsy and neurodegeneration, and can regulate neuronal excitability through interactions with ion channels2. Polyamines have been linked to pain, showing altered levels in human persistent pain states and modulation of pain behaviour in animal models3. However, the systems governing polyamine transport within the nervous system remain unclear. Here, undertaking a genome-wide association study (GWAS) of chronic pain intensity in the UK Biobank (UKB), we found a significant association between pain intensity and variants mapping to the SLC45A4 gene locus. In the mouse nervous system, Slc45a4 expression is enriched in all sensory neuron subtypes within the dorsal root ganglion, including nociceptors. Cell-based assays show that SLC45A4 is a selective plasma membrane polyamine transporter, and the cryo-electron microscopy (cryo-EM) structure reveals a regulatory domain and basis for polyamine recognition. Mice lacking SLC45A4 show normal mechanosensitivity but reduced sensitivity to noxious heat- and algogen-induced tonic pain that is associated with reduced excitability of C-polymodal nociceptors. Our findings therefore establish a role for neuronal polyamine transport in pain perception and identify a target for therapeutic intervention in pain treatment.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: D.L.B. has acted as a consultant for 5AM Ventures, AditumBio, AstraZeneca, Biogen, Biointervene, Combigene, GSK, Ionis, Lexicon Therapeutics, Neuvati, Novo Ventures, Olipass, Orion, Replay, SC Health Managers, Third Rock Ventures, Vida Ventures, Vertexon on behalf of Oxford University Innovation over the past two years. The PAINSTORM consortium received funding from Eli Lilly and AstraZeneca. The DOLORisk consortium received funding from Eli Lilly. J.E.L. is an employee of AstraZeneca. H.R.K. is a member of advisory boards for Altimmune and Clearmind Medicine; a consultant to Sobrera Pharmaceuticals and Altimmune; the recipient of research funding and medication supplies for an investigator-initiated study from Alkermes; a member of the American Society of Clinical Psychopharmacology’s Alcohol Clinical Trials Initiative, which was supported in the past three years by Alkermes, Dicerna, Ethypharm, Imbrium, Indivior, Kinnov, Lilly, Otsuka and Pear Therapeutics; and is listed as an inventor on US provisional patent ‘Multi-ancestry Genome-wide Association Meta-analysis of Buprenorphine Treatment Response’.
Figures














Update of
-
Identification of SLC45A4 as a pain gene encoding a neuronal polyamine transporter.bioRxiv [Preprint]. 2024 Sep 30:2024.09.25.615046. doi: 10.1101/2024.09.25.615046. bioRxiv. 2024. Update in: Nature. 2025 Oct;646(8084):404-412. doi: 10.1038/s41586-025-09326-y. PMID: 41031034 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
