

Functional characterization of the Csm1-like protein TITAN 9 in Arabidopsis thaliana
- PMID: 40837229
- PMCID: PMC12362390
- DOI: 10.1016/j.isci.2025.113251
Functional characterization of the Csm1-like protein TITAN 9 in Arabidopsis thaliana
Abstract
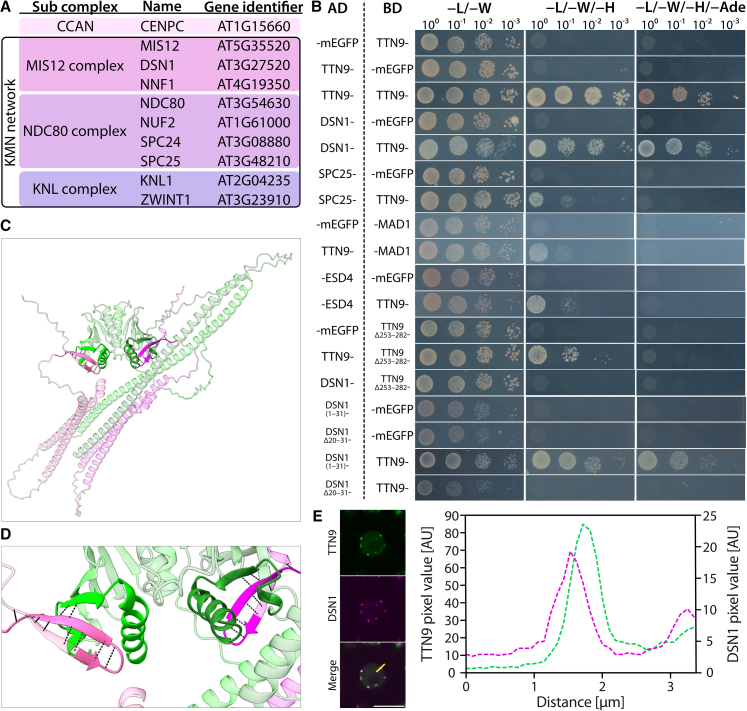
Kinetochores are essential for chromosome segregation in eukaryotes. An important component of kinetochores in opisthokonta is Csm1. However, its function appears to be diversified and, while Csm1 in budding yeast is a component of the monopolin complex mediating mono-orientation of sister kinetochores during meiosis I, the fission yeast homolog Pcs1 prevents merotelic spindle microtubule attachments during mitosis and meiosis II. Here, we have characterized TITAN9 (TTN9), a distantly related Csm1-like protein in the flowering plant Arabidopsis. TTN9 accumulates in mitotic and meiotic tissue and localizes to centromeres throughout the cell cycle. By analyzing proteome-wide TTN9 associated proteins, we identified a substantial subset of the Arabidopsis kinetochore proteome, including DSN1, mirroring known Csm1 interactions in yeast. While homozygous ttn9 mutants are not viable, a meiosis-specific knock-down of TTN9 causes chromosome segregation defects and split centromeres during meiosis I. These findings suggest that Csm1-like proteins contribute to conserved kinetochore functions across eukaryotes.
Keywords: plant Biology; plant bioinformatics; plant development; plant evolution; plant genetics; plant physiology; plant systematics.
© 2025 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Chelysheva L., Diallo S., Vezon D., Gendrot G., Vrielynck N., Belcram K., Rocques N., Márquez-Lema A., Bhatt A.M., Horlow C., et al. AtREC8 and AtSCC3 are essential to the monopolar orientation of the kinetochores during meiosis. J. Cell Sci. 2005;118:4621–4632. - PubMed
-
- Yokobayashi S., Watanabe Y. The kinetochore protein Moa1 enables cohesion-mediated monopolar attachment at meiosis I. Cell. 2005;123:803–817. - PubMed
-
- Watanabe Y., Nurse P. Cohesin Rec8 is required for reductional chromosome segregation at meiosis. Nature. 1999;400:461–464. - PubMed
-
- Sakuno T., Tada K., Watanabe Y. Kinetochore geometry defined by cohesion within the centromere. Nature. 2009;458:852–858. - PubMed
LinkOut - more resources
Full Text Sources
