

Honeybee-Gilliamella synergy in carbohydrate metabolism enhances host thermogenesis in cold acclimation
- PMID: 40850953
- PMCID: PMC12375794
- DOI: 10.1038/s41522-025-00798-4
Honeybee-Gilliamella synergy in carbohydrate metabolism enhances host thermogenesis in cold acclimation
Abstract
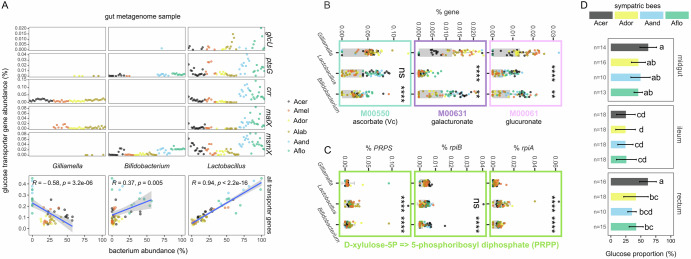
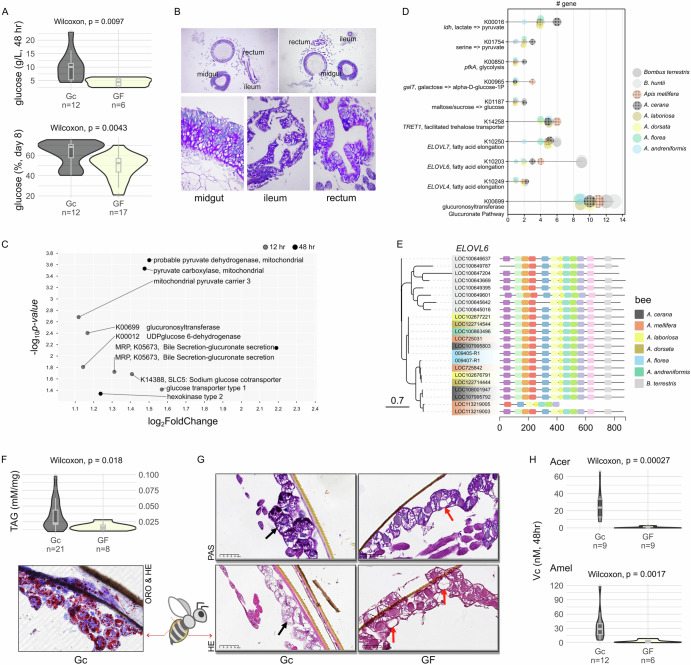
How gut symbionts contribute to host adaptation remains largely elusive. Studying co-diversified honeybees and gut bacteria across climates, we found cold-adapted species (Apis mellifera, A. cerana) exhibit enhanced genomic capacity for glucose, pyruvate, lipid and glucuronate production versus tropical species. Metagenomics revealed Gilliamella as the most enriched gut bacterium in cold-adapted bees. Germ-free honeybees inoculated with the Gilliamella from A. cerana showed increased activity, body temperature and fat storage upon cold exposure. Saccharide metabolomics demonstrated higher hindgut glucose levels in Gilliamella-colonized A. mellifera versus germ-free bees, and in A. cerana versus three sympatric tropical species. Although Gilliamella can hydrolyze β-glucan into glucose, cultural experiments suggest it preferentially degrades glucuronate to pyruvate. In turn, monocolonized bees upregulated hindgut glucose/pyruvate utilization while increasing glucuronate provision, suggesting nutritional complementarity. Gilliamella's transporter genes predominantly target ascorbate (a glucuronate derivative), which is elevated in inoculated hindguts. Accordingly, Gilliamella converts ascorbate to D-xylulose-5P (promoting lipogenesis), while showing reduced growth on glucuronate/ascorbate versus glucose, potentially minimizing glucose competition with hosts. We revealed a highly coordinated host-symbiont metabolic synergy enhancing host energy acquisition for cold adaptation.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures




References
MeSH terms
Substances
Grants and funding
- RDF-23-01-067/Research Development Fund of Xi'an Jiaotong-Liverpool University
- 2023YFD2201804/Program of the Ministry of Science and Technology of China
- 2023YFD2201804/Program of the Ministry of Science and Technology of China
- 32470455/National Natural Science Foundation of China
- 2023IOZ0104/program from Institute of Zoology, Chinese Academy of Sciences
LinkOut - more resources
Full Text Sources
