

Cholecystokinin-expressing GABA neurons elicit long-term potentiation in the cortical inhibitory synapses and attenuate sound-shock associative memory
- PMID: 40854983
- PMCID: PMC12379247
- DOI: 10.1038/s41598-025-17065-3
Cholecystokinin-expressing GABA neurons elicit long-term potentiation in the cortical inhibitory synapses and attenuate sound-shock associative memory
Abstract
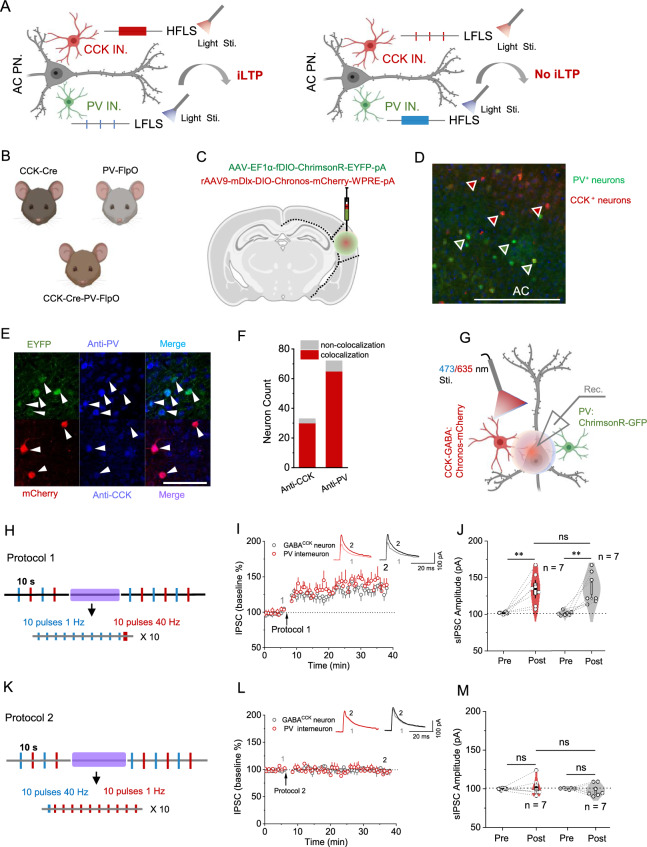
Neuronal interactions between inhibitory and excitatory neurons play a pivotal role in regulating the balance of excitation and inhibition in the central nervous system (CNS). Consequently, the efficacy of inhibitory/excitatory synapses profoundly affects neural network processing and overall neuronal functions. Here, we describe a novel form of long-term potentiation (LTP) induced at cortical inhibitory synapses and its behavioral consequences. We show that high-frequency laser stimulation (HFLS) of GABAergic neurons elicit inhibitory LTP (i-LTP) in pyramidal neurons of the auditory cortex (AC). The selective activation of cholecystokinin-expressing GABA (GABACCK) neurons is essential for the formation of HFLS-induced i-LTP, rather than the classical parvalbumin (PV) neurons and somatostatin (SST) neurons. Intriguingly, i-LTP can be evoked in the AC by adding the exogenous neuropeptide CCK when PV neurons and SST neurons are selectively activated in PV-Cre and SST-Cre mice, respectively. Additionally, we discovered that low-frequency laser stimulation (LFLS) of PV neurons paired with HFLS of GABACCK neurons potentiates the inhibitory effect of PV interneurons on pyramidal neurons, thereby generating heterosynaptic i-LTP in the AC. Notably, light activation of GABACCK neurons in CCK-Cre mice significantly attenuates sound- shock associative memory, while stimulation of PV neurons does not affect this memory in PV-Cre mice. In conclusion, these results demonstrate a critical mechanism regulating the excitation-inhibition balance and modulating learning and memory in cortical circuits. This mechanism might serve as a potential target for the treatment of neurological disorders, including epilepsy and Alzheimer's disease.
Keywords: Auditory cortex; Cholecystokinin; Interneuron; Long-term potentiation; Neurological disorders.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Competing interests: The authors declare no competing interests.
Figures





References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
