

Single-molecule fluorescence reveals the DNA unwinding mechanism of mitochondrial helicase TWINKLE and its interplay with single-stranded DNA-binding proteins
- PMID: 40867054
- PMCID: PMC12390753
- DOI: 10.1093/nar/gkaf803
Single-molecule fluorescence reveals the DNA unwinding mechanism of mitochondrial helicase TWINKLE and its interplay with single-stranded DNA-binding proteins
Abstract
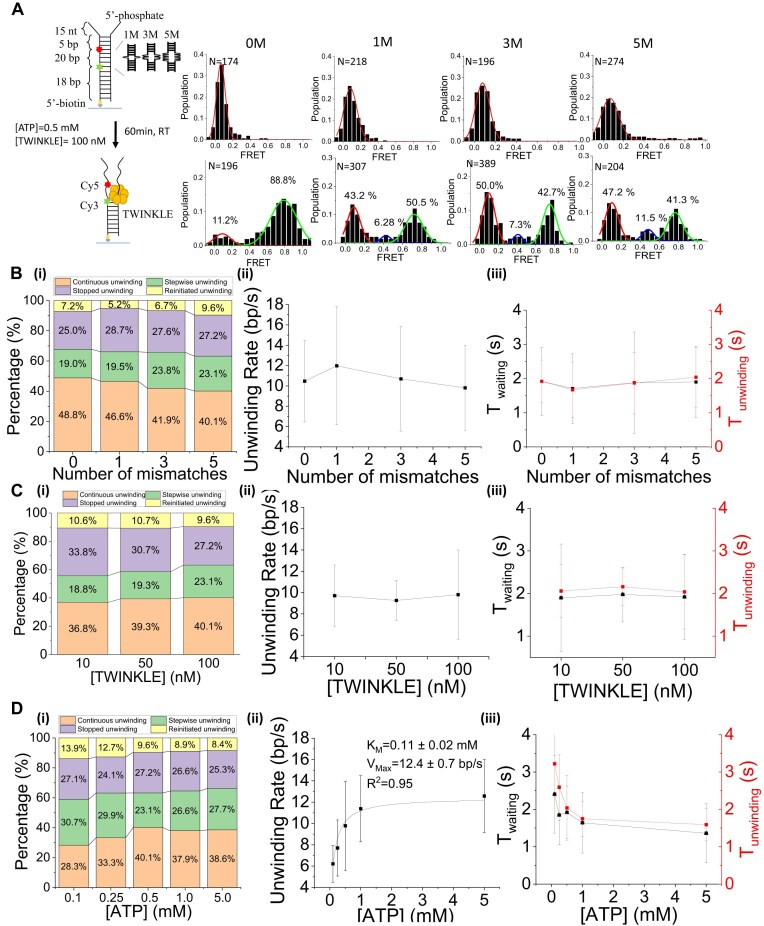
The mitochondrial DNA helicase TWINKLE, a hexameric ring-shaped helicase, plays a crucial role in maintaining mitochondrial DNA integrity. TWINKLE translocates along one DNA strand, unwinding the duplex by excluding the complementary strand through coordinated ATP hydrolysis. However, the precise mechanisms underlying this process remain incompletely understood. In this study, we utilized single-molecule Förster Resonance Energy Transfer (smFRET) to investigate the mechanisms of TWINKLE-mediated DNA unwinding. Our results reveal that TWINKLE occasionally pauses during unwinding, with the rate of unwinding and the duration of pausing strongly influenced by ATP concentration, but not by the presence of DNA mismatches or mitochondrial single-stranded DNA-binding protein (mtSSB). These findings suggest that the pausing events primarily arise from stochastic ATP hydrolysis within the helicase subunits. DNA mismatches exacerbate TWINKLE's pausing and dissociation from DNA, thereby impairing DNA unwinding. In contrast, mtSSB significantly mitigates helicase dissociation by stabilizing TWINKLE-DNA interactions. This study provides novel insights into the functional dynamics of TWINKLE, highlighting the role of ATP hydrolysis in orchestrating single-stranded DNA translocation, the detrimental effects of DNA mismatches on DNA unwinding, and the critical role of mtSSB in supporting helicase function.
© The Author(s) 2025. Published by Oxford University Press.
Conflict of interest statement
None declared.
Figures






Similar articles
-
Autoregulation of the real-time kinetics of the human mitochondrial replicative helicase.Nat Commun. 2025 Jul 1;16(1):5460. doi: 10.1038/s41467-025-60289-0. Nat Commun. 2025. PMID: 40592829 Free PMC article.
-
ATPase-dependent duplex nucleic acid unwinding by SARS-CoV-2 nsP13 relies on facile binding and translocation along single-stranded nucleic acid.J Biol Chem. 2025 Jul;301(7):110373. doi: 10.1016/j.jbc.2025.110373. Epub 2025 Jun 12. J Biol Chem. 2025. PMID: 40516869 Free PMC article.
-
Stimulation of Variant Forms of the Mitochondrial DNA Helicase Twinkle by the Mitochondrial Single-Stranded DNA-Binding Protein.Methods Mol Biol. 2021;2281:313-322. doi: 10.1007/978-1-0716-1290-3_20. Methods Mol Biol. 2021. PMID: 33847968
-
Chromatin gatekeeper and modifier CHD proteins in development, and in autism and other neurological disorders.Psychiatr Genet. 2023 Dec 1;33(6):213-232. doi: 10.1097/YPG.0000000000000353. Epub 2023 Oct 16. Psychiatr Genet. 2023. PMID: 37851134 Review.
-
TWINKLE and Other Human Mitochondrial DNA Helicases: Structure, Function and Disease.Genes (Basel). 2020 Apr 9;11(4):408. doi: 10.3390/genes11040408. Genes (Basel). 2020. PMID: 32283748 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
