

Repurposing the Antibiotic D-Cycloserine for the Treatment of Hyperpigmentation: Therapeutic Potential and Mechanistic Insights
- PMID: 40869042
- PMCID: PMC12386294
- DOI: 10.3390/ijms26167721
Repurposing the Antibiotic D-Cycloserine for the Treatment of Hyperpigmentation: Therapeutic Potential and Mechanistic Insights
Abstract
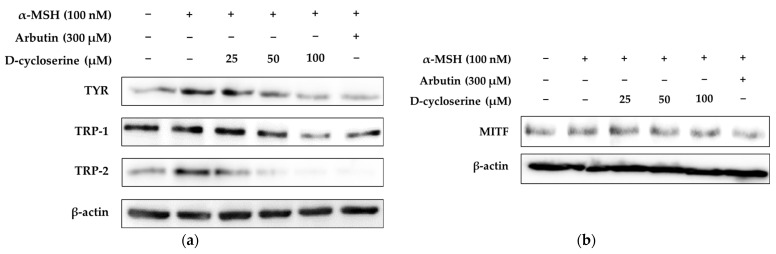
Melanin overproduction contributes to hyperpigmentation disorders such as melasma and solar lentigines, leading to increasing demand for safe and effective skin-lightening agents. D-cycloserine (DCS), a known antimicrobial agent, has not been previously evaluated for dermatological applications. This study aimed to explore the potential of DCS as a novel anti-melanogenic compound and to elucidate its underlying molecular mechanisms in melanogenesis inhibition. The cytotoxicity and anti-melanogenic effects of DCS were assessed in B16F10 melanoma cells stimulated with α-MSH. Cell viability was determined via MTT assays, while melanin content, tyrosinase activity, and the expression levels of MITF, TYR, TRP-1, TRP-2, and major signaling proteins (e.g., CREB, MAPKs, GSK-3β/β-catenin) were evaluated using colorimetric assays and Western blotting. A 3D human skin model was also used to confirm in vitro findings, and a primary skin irritation test was conducted to assess dermal safety. DCS significantly reduced α-MSH-induced melanin content and tyrosinase activity without cytotoxicity at concentrations ≤100 µM. It downregulated MITF and melanogenic enzyme expression and modulated signaling pathways by enhancing ERK activation while inhibiting CREB, JNK, and p38 phosphorylation. Additionally, DCS suppressed β-catenin stabilization via GSK-3β activation. These effects were confirmed in a 3D human skin model, and a clinical skin irritation study revealed no adverse reactions in human volunteers. DCS exerts its anti-melanogenic effect by targeting multiple pathways, including CREB/MITF, MAPK, and GSK-3β/β-catenin signaling. Its efficacy and safety profiles support its potential as a novel cosmeceutical agent for the treatment of hyperpigmentation. Further clinical studies are warranted to confirm its therapeutic utility in human skin pigmentation disorders.
Keywords: B16F10; CREB; D-cycloserine; MAPK pathway; PI3K/Akt pathway; cosmeceutical; drug repurposing; hyperpigmentation; melanogenesis; β-catenin.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures







References
-
- Akl J., Lee S., Ju H.J., Parisi R., Kim J.Y., Jeon J.J., Heo Y.W., Eleftheriadou V., Hamzavi I., Griffiths C.E.M., et al. Global Vitiligo Atlas. Estimating the burden of vitiligo: A systematic review and modelling study. Lancet Public Health. 2024;9:e386–e396. doi: 10.1016/S2468-2667(24)00026-4. - DOI - PubMed
-
- Pinzi L., Bisi N., Rastelli G. How drug repurposing can advance drug discovery: Challenges and opportunities. Front. Drug Discov. 2024;4:1460100. doi: 10.3389/fddsv.2024.1460100. - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
