

The deacetylases HDAC1/HDAC2 control JAK2V617F-STAT signaling through the ubiquitin ligase SIAH2
- PMID: 40877230
- PMCID: PMC12394589
- DOI: 10.1038/s41392-025-02369-7
The deacetylases HDAC1/HDAC2 control JAK2V617F-STAT signaling through the ubiquitin ligase SIAH2
Abstract
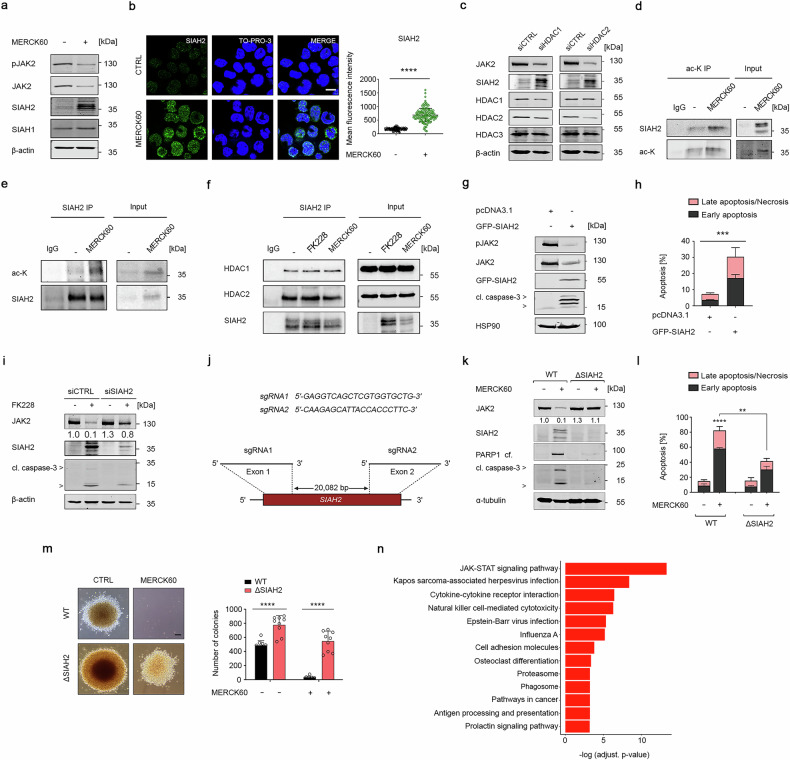
Epigenetic modulators of the histone deacetylase (HDAC) family control key biological processes and are frequently dysregulated in cancer. There is superior activity of HDAC inhibitors (HDACi) in patients with myeloproliferative neoplasms (MPNs) that carry the Janus kinase-2 point mutant JAK2V617F. This constitutively active tyrosine kinase activates signal-transducer-and-activator-of-transcription (STAT) transcription factors to promote cell proliferation and inflammatory processes. We reveal that the inhibition of HDAC1/HDAC2 with the clinically advanced HDACi romidepsin, the experimental HDACi entinostat and MERCK60, and genetic depletion of HDAC1/HDAC2 induce apoptosis and long-term growth arrest of primary and permanent MPN cells in vitro and in vivo. This treatment spares normal hematopoietic stem cells and does not compromise blood cell differentiation. At the molecular level, HDAC1 and HDAC2 control the protein stability of SIAH2 through acetylation. Genetic knockout experiments show that SIAH2 accelerates the proteasomal degradation of JAK2V617F in conjunction with the E2 ubiquitin-conjugating enzyme UBCH8. SIAH2 binds to the surface-exposed SIAH degron motif VLP1002 in the catalytic domain of JAK2V617F. At the functional level, SIAH2 knockout MPN cells are significantly less sensitive to HDACi. Global RNA sequencing verifies that JAK-STAT signaling is a prime target of SIAH2. Moreover, HDAC1 is an adverse prognostic factor in patients with acute myeloid leukemia (n = 150, p = 0.02), being a possible complication of MPNs. These insights reveal a previously unappreciated link between HDAC1/HDAC2 as key molecular targets, the still undefined regulation of cytoplasmic-to-nuclear signaling by HDACs, and how HDACi kill JAK2V617F-positive cells from MPN patients and mice with JAK2V617F in vitro and in vivo.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: O.H.K. declares paid consultant activity for BASF, Ludwigshafen, Germany, and the patents “The use of molecular markers for the preclinical and clinical profiling of inhibitors of enzymes having histone deacetylase activity”, “Novel HDAC6 inhibitors and their uses”, and “Synthesis, pharmacology and use of new and selective FMS-like tyrosine kinase 3 (FLT3) inhibitors”. The BASF did not influence the study, and its products are not discussed in this work. All the other authors declare no conflict of interests.
Figures




References
-
- Mustafa, A. M. & Krämer, O. H. Pharmacological modulation of the crosstalk between aberrant janus kinase signaling and epigenetic modifiers of the histone deacetylase family to treat cancer. Pharm. Rev.75, 35–61 (2023). - PubMed
-
- Jayavelu, A. K. et al. Splicing factor YBX1 mediates persistence of JAK2-mutated neoplasms. Nature588, 157–163 (2020). - PubMed
MeSH terms
Substances
Grants and funding
- KR2291/Deutsche Forschungsgemeinschaft (German Research Foundation)
- 329045328/Deutsche Forschungsgemeinschaft (German Research Foundation)
- 318346496/Deutsche Forschungsgemeinschaft (German Research Foundation)
- 517204983/Deutsche Forschungsgemeinschaft (German Research Foundation)
- 320028127/Deutsche Forschungsgemeinschaft (German Research Foundation)
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
