

Identification and replication of sex-dimorphic protein quantitative trait loci across multiple ancestries and their associations with diseases
- PMID: 40877285
- PMCID: PMC12394655
- DOI: 10.1038/s41598-025-10031-z
Identification and replication of sex-dimorphic protein quantitative trait loci across multiple ancestries and their associations with diseases
Abstract
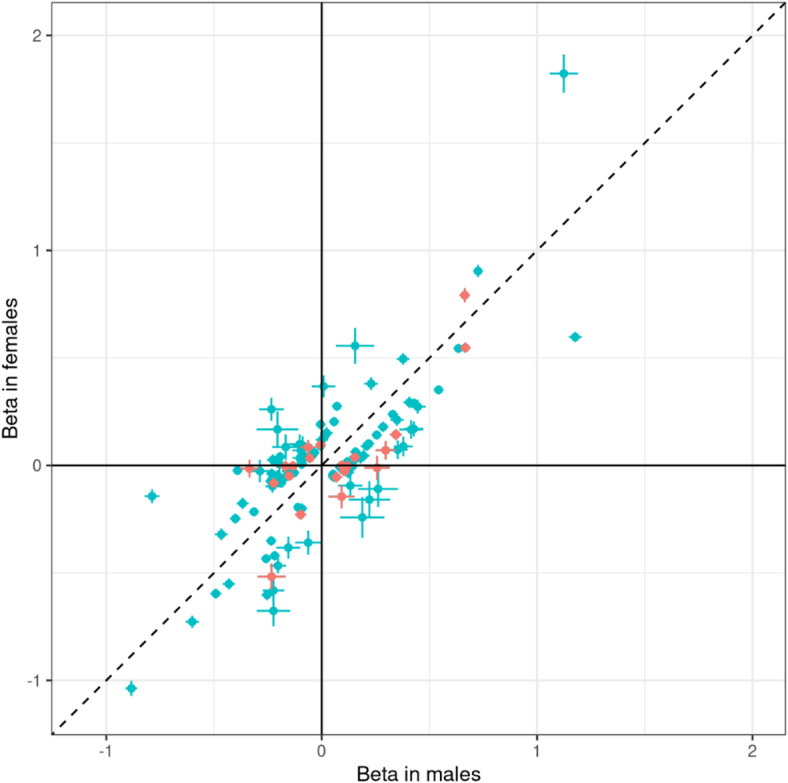
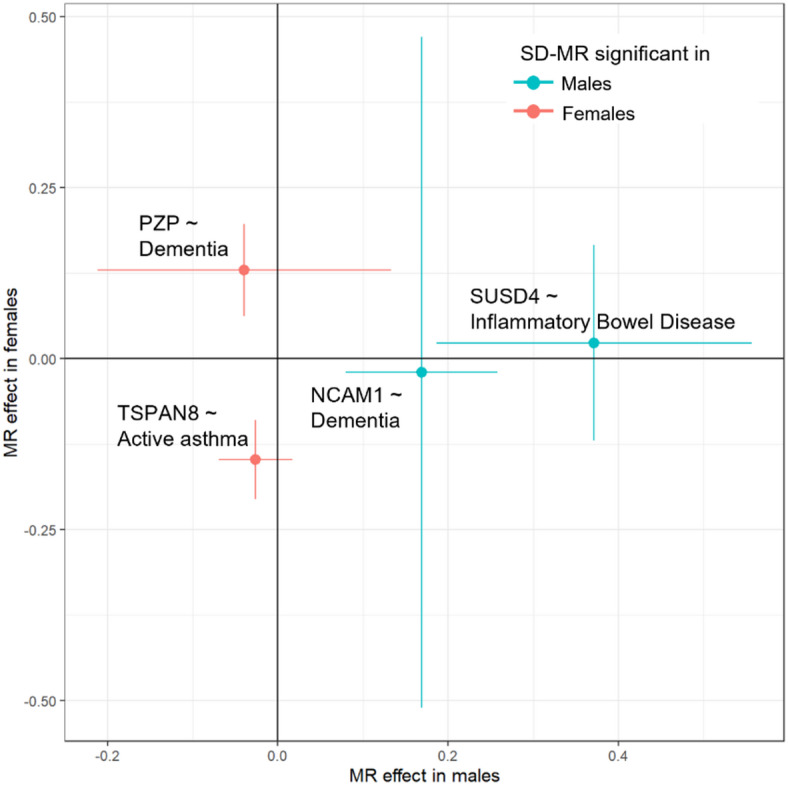
Males and females exhibit differences in proteome profiles associated with disease risk. However, sex-dimorphic protein quantitative trait loci (SD-pQTL) and their effects on sex differences in health disorders have not been thoroughly investigated. We conducted a sex-stratified, genome-wide association study on 2,922 proteins using data from 30,272 individuals of Caucasian ancestry from the UK Biobank and compared the estimated effects on protein levels of these variants in the men and women to identify SD-pQTLs. The identified SD-pQTLs were replicated using data from two Japanese cohorts (comprising 2,886 and 1,394 individuals, respectively), as well as from 1,990 Finnish, 630 South Asian, and 662 Black ancestry individuals. Sex-dimorphic pleiotropy and the causal relationship between protein levels and health disorders were assessed using the identified SD-pQTLs. We identified 113 SD-pQTLs associated with 65 proteins. Of the 113 SD-pQTLs, 52 were significant in both sexes, five were not significant in either sex, and 42 and 14 were significant only in males and females, respectively. Variant rs2270416 was significantly associated with the CDH15 protein in both sexes but showed opposite effect direction in men and women. Of the 113 SD-pQTLs identified, a total of 41 were replicated in a meta-analysis encompassing Japanese, South Asian, and Black ancestry individuals. SD-pQTLs for proteins APOE (rs157581) and SNAP25 (rs4420638) exhibited sex-dimorphic associations with dementia, indicating sex dimorphic pleiotropy in both proteins and health disorders. From sex-stratified Mendelian randomization using the SD-pQTLs, proteins NCAM1 and PZP showed significant causal relationship with dementia in males and females, respectively. The present study provides evidence of sex-dimorphic genetic architecture in protein-level regulation, elucidating the proteo-genetic architecture for sex differences in human variation.
Keywords: Proteome profiles; Sex differences; Sex-dimorphic protein quantitative trait loci.
© 2025. Crown.
Conflict of interest statement
Competing interests: The authors declare no competing interests. Ethics approval and consent to participants: The UK Biobank project was approved by the National Research Ethics Service Committee North West-Haydock (REC reference: 11/NW/0382). Participants provided written informed consent to participate in the UK Biobank. An electronic signed consent was obtained from the participants. This research was conducted using the UK Biobank Resource under project 44986. This work uses data provided by patients and collected by the NHS as part of their care and support. BioBank Japan project was approved by the Institutional Review Board of the University of Tokyo (2023-77-0118 and 2022-61-0119). Informed consent was obtained from all participants. The JCTF was approved by the ethical committees of Keio University School of Medicine, Osaka University Graduate School of Medicine, and affiliated institutes. Informed consent was obtained from all participants. Study subjects in FinnGen provided informed consent for biobank research, based on the Finnish Biobank Act. Alternatively, separate research cohorts, collected prior the Finnish Biobank Act came into effect (in September 2013) and start of FinnGen (August 2017), were collected based on study-specific consents and later transferred to the Finnish biobanks after approval by Fimea (Finnish Medicines Agency), the National Supervisory Authority for Welfare and Health. Recruitment protocols followed the biobank protocols approved by Fimea. The Coordinating Ethics Committee of the Hospital District of Helsinki and Uusimaa (HUS) statement number for the FinnGen study is Nr HUS/990/2017. The FinnGen study is approved by Finnish Institute for Health and Welfare (permit numbers: THL/2031/6.02.00/2017, THL/1101/5.05.00/2017, THL/341/6.02.00/2018, THL/2222/6.02.00/2018, THL/283/6.02.00/2019, THL/1721/5.05.00/2019 and THL/1524/5.05.00/2020), Digital and population data service agency (permit numbers: VRK43431/2017-3, VRK/6909/2018-3, VRK/4415/2019-3), the Social Insurance Institution (permit numbers: KELA 58/522/2017, KELA 131/522/2018, KELA 70/522/2019, KELA 98/522/2019, KELA 134/522/2019, KELA 138/522/2019, KELA 2/522/2020, KELA 16/522/2020), Findata permit numbers THL/2364/14.02/2020, THL/4055/14.06.00/2020, THL/3433/14.06.00/2020, THL/4432/14.06/2020, THL/5189/14.06/2020, THL/5894/14.06.00/2020, THL/6619/14.06.00/2020, THL/209/14.06.00/2021, THL/688/14.06.00/2021, THL/1284/14.06.00/2021, THL/1965/14.06.00/2021, THL/5546/14.02.00/2020, THL/2658/14.06.00/2021, THL/4235/14.06.00/2021, Statistics Finland (permit numbers: TK-53-1041-17 and TK/143/07.03.00/2020 (earlier TK-53-90-20) TK/1735/07.03.00/2021, TK/3112/07.03.00/2021) and Finnish Registry for Kidney Diseases permission/extract from the meeting minutes on 4th July 2019. The Biobank Access Decisions for FinnGen samples and data utilized in FinnGen Data Freeze 12 include: THL Biobank BB2017_55, BB2017_111, BB2018_19, BB_2018_34, BB_2018_67, BB2018_71, BB2019_7, BB2019_8, BB2019_26, BB2020_1, BB2021_65, Finnish Red Cross Blood Service Biobank 7.12.2017, Helsinki Biobank HUS/359/2017, HUS/248/2020, HUS/430/2021 § 28, § 29, HUS/150/2022 § 12, § 13, § 14, § 15, § 16, § 17, § 18, § 23, § 58, § 59, HUS/128/2023 § 18, Auria Biobank AB17-5154 and amendment #1 (August 17 2020) and amendments BB_2021 − 0140, BB_2021 − 0156 (August 26 2021, Feb 2 2022), BB_2021 − 0169, BB_2021 − 0179, BB_2021 − 0161, AB20-5926 and amendment #1 (April 23 2020) and it´s modifications (Sep 22 2021), BB_2022 − 0262, BB_2022 − 0256, Biobank Borealis of Northern Finland_2017_1013, 2021_5010, 2021_5010 Amendment, 2021_5018, 2021_5018 Amendment, 2021_5015, 2021_5015 Amendment, 2021_5015 Amendment_2, 2021_5023, 2021_5023 Amendment, 2021_5023 Amendment_2, 2021_5017, 2021_5017 Amendment, 2022_6001, 2022_6001 Amendment, 2022_6006 Amendment, 2022_6006 Amendment, 2022_6006 Amendment_2, BB22-0067, 2022_0262, 2022_0262 Amendment, Biobank of Eastern Finland 1186/2018 and amendment 22§/2020, 53§/2021, 13§/2022, 14§/2022, 15§/2022, 27§/2022, 28§/2022, 29§/2022, 33§/2022, 35§/2022, 36§/2022, 37§/2022, 39§/2022, 7§/2023, 32§/2023, 33§/2023, 34§/2023, 35§/2023, 36§/2023, 37§/2023, 38§/2023, 39§/2023, 40§/2023, 41§/2023, Finnish Clinical Biobank Tampere MH0004 and amendments (21.02.2020 & 06.10.2020), BB2021-0140 8§/2021, 9§/2021, § 9/2022, § 10/2022, § 12/2022, 13§/2022, § 20/2022, § 21/2022, § 22/2022, § 23/2022, 28§/2022, 29§/2022, 30§/2022, 31§/2022, 32§/2022, 38§/2022, 40§/2022, 42§/2022, 1§/2023, Central Finland Biobank 1-2017, BB_2021 − 0161, BB_2021 − 0169, BB_2021 − 0179, BB_2021 − 0170, BB_2022 − 0256, BB_2022 − 0262, BB22-0067, Decision allowing to continue data processing until 31st Aug 2024 for projects: BB_2021 − 0179, BB22-0067,BB_2022 − 0262, BB_2021 − 0170, BB_2021 − 0164, BB_2021 − 0161, and BB_2021 − 0169, and Terveystalo Biobank STB 2018001 and amendment 25th Aug 2020, Finnish Hematological Registry and Clinical Biobank decision 18th June 2021, Arctic biobank P0844: ARC_2021_1001.
Figures

References
-
- Heron, M. P. Deaths: leading causes for 2018 (2021). - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
