

Iminium Catalysis in Natural Diels-Alderase
- PMID: 40881614
- PMCID: PMC12381789
- DOI: 10.1038/s41929-025-01294-w
Iminium Catalysis in Natural Diels-Alderase
Abstract
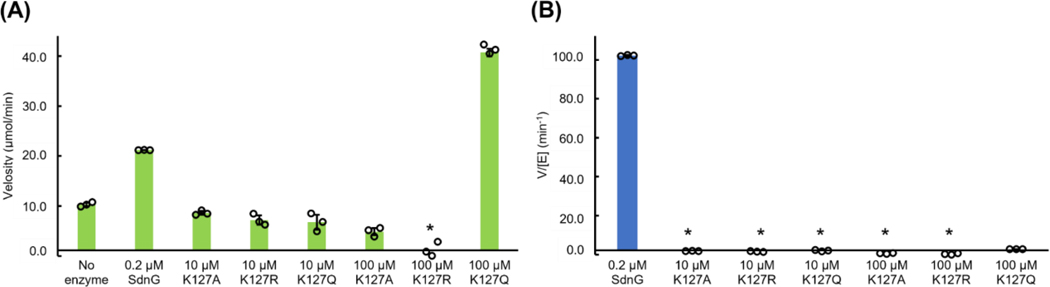
Iminium-catalyzed cycloaddition is one of the most prominent examples of organocatalysis, yet a biological counterpart has not been reported despite the wide-spread occurrence of iminium adducts in enzymes. Here, we present biochemical, structural, and computational evidence for iminium catalysis by the natural Diels-Alderase SdnG that catalyzes norbornene formation in sordarin biosynthesis. A Schiff base adduct between the ε-nitrogen of active site K127 and the aldehyde group of the enal dienophile was revealed by structural analysis and captured under catalytic conditions via borohydride reduction. This Schiff base adduct positions the substrate into near-attack conformation and decreases the transition state barrier of Diels-Alder cyclization by 8.3 kcal/mol via dienophile activation. A hydrogen bond network consisting of a catalytic triad is proposed to facilitate proton transfer required for iminium formation. This work establishes a new mode of catalysis for Diels-Alderases and points the way to the design of novel iminium based (bio)catalysts.
Figures








References
-
- MacMillan DWC The advent and development of organocatalysis. Nature 455, 304–308 (2008). - PubMed
-
- Ahrendt KA, Borths CJ & MacMillan DWC New Strategies for Organic Catalysis: The First Highly Enantioselective Organocatalytic Diels−Alder Reaction. J. Am. Chem. Soc 122, 4243–4244 (2000).
-
- Gefflaut T, Blonski C, Perie J. & Willson M. Class I aldolases: Substrate specificity, mechanism, inhibitors and structural aspects. Prog. Biophys. Mol. Biol 63, 301–340 (1995). - PubMed
-
- Piersen CE, McCullough AK & Lloyd RS AP lyases and dRPases: commonality of mechanism. Mutat. Res. Repair 459, 43–53 (2000). - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources