

β-Amyloid induces microglial expression of GPC4 and APOE leading to increased neuronal tau pathology and toxicity
- PMID: 40883746
- PMCID: PMC12398132
- DOI: 10.1186/s13024-025-00883-4
β-Amyloid induces microglial expression of GPC4 and APOE leading to increased neuronal tau pathology and toxicity
Abstract
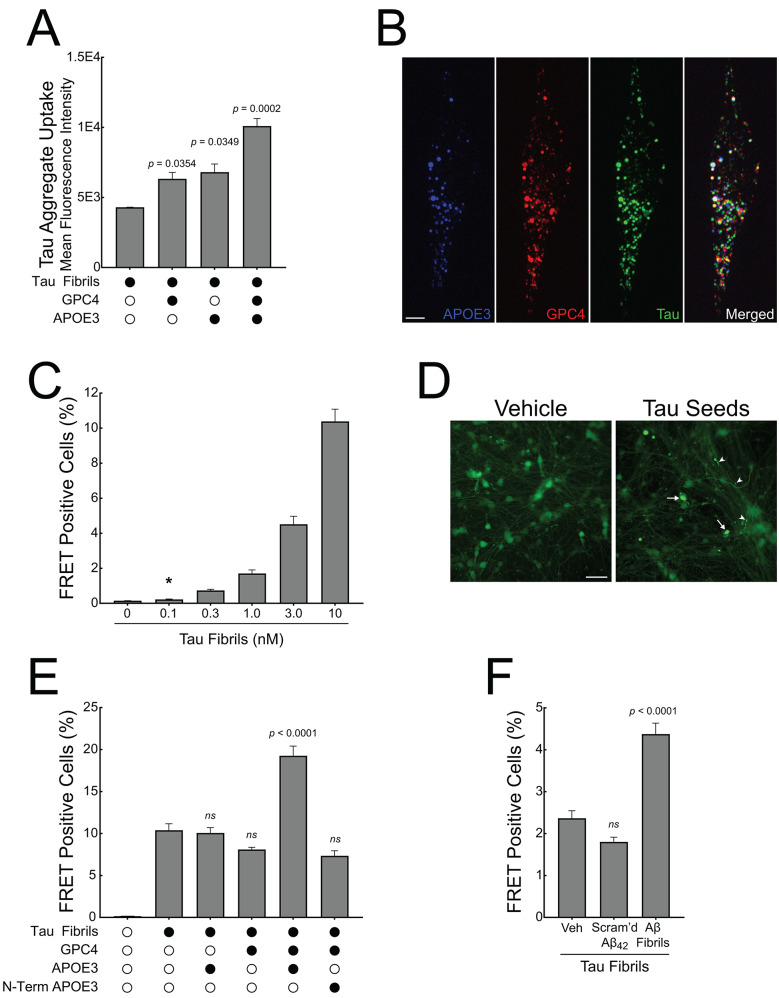
To define how Aβ pathology alters microglia function in Alzheimer’s disease, we profiled the microglia surfaceome following treatment with Aβ fibrils. Our findings reveal that Aβ-associated human microglia upregulate Glypican 4 (GPC4), a GPI-anchored heparan sulfate proteoglycan (HSPG). Glial GPC4 expression exacerbates motor deficits and reduces lifespan in a Drosophila amyloidosis model, implicating GPC4 in a toxic neurodegenerative program. In cell culture, GPC4 enhances microglia phagocytosis of tau aggregates, and shed GPC4 can act in trans to facilitate tau aggregate uptake and seeding in neurons. Additionally, our data demonstrate that GPC4-mediated effects are amplified in the presence of APOE. In human Alzheimer’s disease brain, microglial GPC4 expression surrounding Aβ plaques correlates with neuritic tau pathology, supporting a pathological link between amyloid, GPC4, and tau. These studies define a mechanistic pathway by which Aβ primes microglia to promote tau pathology via HSPGs and APOE.
Supplementary Information: The online version contains supplementary material available at 10.1186/s13024-025-00883-4.
Keywords: Drosophila model; APOE; Alzheimer’s disease; Amyloid; Astrocytes; GPC4; Heparan sulfate proteoglycans; Microglia; Neuroinflammation; Tau seeding.
Conflict of interest statement
Declarations. Ethics approval and consent to participate: All animal procedures were conducted in accordance with the guidelines set forth by the Institutional Animal Care and Use Committee at the University of Virginia School of Medicine, and study protocols were reviewed and approved prior to performing the experimental procedures described. Consent for publication: All authors have approved the contents of this manuscript and provided consent for publication. Competing interests: B.B.H has the following US patents: Therapeutic Anti-Tau Antibodies (US9834596), Method for Detection of Aggregates in Biological Samples (US9910048) and Methods and Compositions Related to Heparinoids (WO2021142312A1). M.K. is a co-scientific founder of Montara Therapeutics and serves on the Scientific Advisory Boards of Engine Biosciences, Casma Therapeutics, Alector, and Montara Therapeutics, and is an advisor to Modulo Bio and Recursion Therapeutics.
Figures







Update of
-
β-Amyloid Induces Microglial Expression of GPC4 and APOE Leading to Increased Neuronal Tau Pathology and Toxicity.bioRxiv [Preprint]. 2025 Feb 25:2025.02.20.637701. doi: 10.1101/2025.02.20.637701. bioRxiv. 2025. Update in: Mol Neurodegener. 2025 Aug 29;20(1):96. doi: 10.1186/s13024-025-00883-4. PMID: 40060520 Free PMC article. Updated. Preprint.
References
-
- Götz J, Chen F, van Dorpe J, Nitsch RM. Formation of neurofibrillary tangles in P301L Tau Transgenic mice induced by Aβ42 fibrils. Science. 2001;293(5534):1491–5. - PubMed
-
- Lewis J, Dickson DW, Lin WL, Chisholm L, Corral A, Jones G, et al. Enhanced neurofibrillary degeneration in Transgenic mice expressing mutant Tau and APP. Science. 2001;293(5534):1487–91. - PubMed
Grants and funding
- R35 GM122451/GM/NIGMS NIH HHS/United States
- P30AG062422/NH/NIH HHS/United States
- K24 AG053435/AG/NIA NIH HHS/United States
- ZEN-22-969903/ALZ/Alzheimer's Association/United States
- R01 NS121101/NS/NINDS NIH HHS/United States
- P30 AG062422/AG/NIA NIH HHS/United States
- Shenandoah Community Foundation/Shenandoah Community Foundation
- P01 AG019724/AG/NIA NIH HHS/United States
- P01AG019724/NH/NIH HHS/United States
- U01AG057195/NH/NIH HHS/United States
- R01NS121101/NS/NINDS NIH HHS/United States
- RF1AG061874/NH/NIH HHS/United States
- BrightFocus Foundation/BrightFocus Foundation
- R35GM122451/GM/NIGMS NIH HHS/United States
- U01 AG057195/AG/NIA NIH HHS/United States
- Brain Institute, University of Virginia/Brain Institute, University of Virginia
- U19 AG063911/AG/NIA NIH HHS/United States
- RF1 AG061874/AG/NIA NIH HHS/United States
- U19AG063911/NH/NIH HHS/United States
- K08 NS133290/NS/NINDS NIH HHS/United States
- K24AG053435/NH/NIH HHS/United States
- 1K08NS133290/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
