

Single-molecule fluorescence microscopy reveals regulatory mechanisms of MYO7A-driven cargo transport in stereocilia of live inner ear hair cells
- PMID: 40890108
- PMCID: PMC12402077
- DOI: 10.1038/s41467-025-63102-0
Single-molecule fluorescence microscopy reveals regulatory mechanisms of MYO7A-driven cargo transport in stereocilia of live inner ear hair cells
Abstract
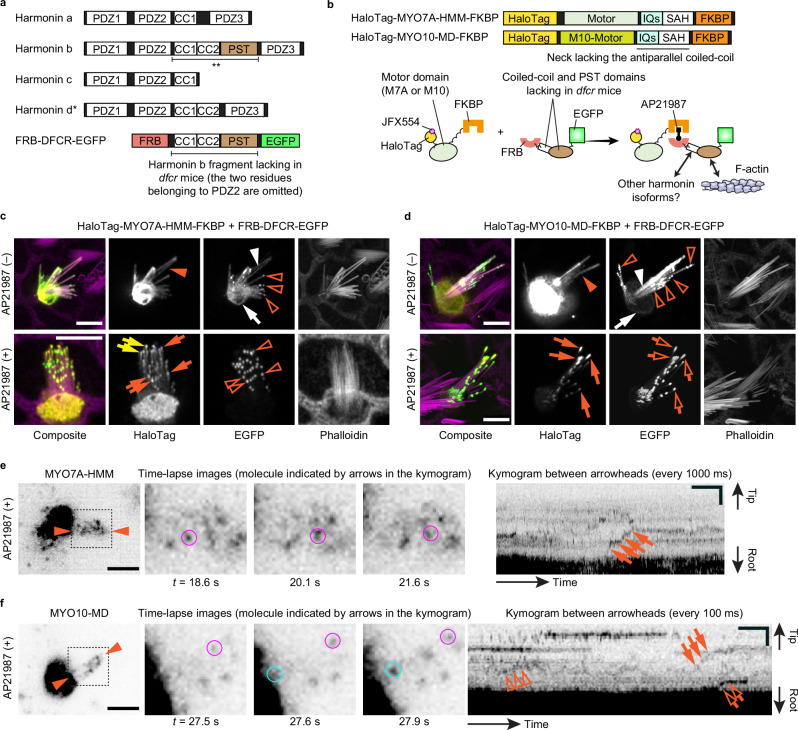
Stereocilia are F-actin-based cylindrical protrusions on the apical surface of inner ear hair cells that function as biological mechanosensors of sound and acceleration. During stereocilia development, specific unconventional myosins transport proteins and phospholipids as cargo and mediate elongation, differentiation and acquisition of the mechanoelectrical transduction (MET). How unconventional myosins localize themselves and cargo in stereocilia using energy from ATP hydrolysis is only partially understood. Here, we developed STELLA-SPIM microscopy to visualize movement of single myosin molecules in live hair cell stereocilia. STELLA-SPIM demonstrated that MYO7A, a component of MET machinery, shows processive movement toward stereocilia tips when chemically dimerized or constitutively activated by missense mutations disabling tail-mediated autoinhibition. Conversely, MYO7A shows step-wise but not processive movement in stereocilia when its tail is tethered to the plasma membrane or F-actin in the presence of MYO7A interacting partners. We posit that MYO7A dimerizes and moves processively in stereocilia when unleashed from autoinhibition.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures





Update of
-
Live-cell single-molecule fluorescence microscopy for protruding organelles reveals regulatory mechanisms of MYO7A-driven cargo transport in stereocilia of inner ear hair cells.bioRxiv [Preprint]. 2024 May 7:2024.05.04.590649. doi: 10.1101/2024.05.04.590649. bioRxiv. 2024. Update in: Nat Commun. 2025 Sep 1;16(1):8149. doi: 10.1038/s41467-025-63102-0. PMID: 38766013 Free PMC article. Updated. Preprint.
-
Live-cell single-molecule fluorescence microscopy for protruding organelles reveals regulatory mechanisms of MYO7A-driven cargo transport in stereocilia of inner ear hair cells.Res Sq [Preprint]. 2024 May 21:rs.3.rs-4369958. doi: 10.21203/rs.3.rs-4369958/v1. Res Sq. 2024. Update in: Nat Commun. 2025 Sep 1;16(1):8149. doi: 10.1038/s41467-025-63102-0. PMID: 38826223 Free PMC article. Updated. Preprint.
References
-
- Tanna, R. J., Lin, J. W. & De Jesus, O. Sensorineural Hearing Loss, <https://www.ncbi.nlm.nih.gov/pubmed/33351419> (2024). - PubMed
-
- Eatock, R. A. & Lysakowski, A. In Vertebrate Hair Cells (eds Ruth A. E., Fay R. R., & Popper A. N.) 348–442 (Springer New York, 2006).
MeSH terms
Substances
Grants and funding
- T32 DC000039/DC/NIDCD NIH HHS/United States
- R00 DC019949/DC/NIDCD NIH HHS/United States
- Z01 DC000039/ImNIH/Intramural NIH HHS/United States
- 1R00DC019949/U.S. Department of Health & Human Services | NIH | National Institute on Deafness and Other Communication Disorders (NIDCD)
- DC000039/U.S. Department of Health & Human Services | NIH | National Institute on Deafness and Other Communication Disorders (NIDCD)
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
