

Mechanistic insights into the T6SS of multi-drug-resistant Aeromonas hydrophila and its role in competition and pathogenesis
- PMID: 40893974
- PMCID: PMC12396204
- DOI: 10.1002/mlf2.70018
Mechanistic insights into the T6SS of multi-drug-resistant Aeromonas hydrophila and its role in competition and pathogenesis
Abstract
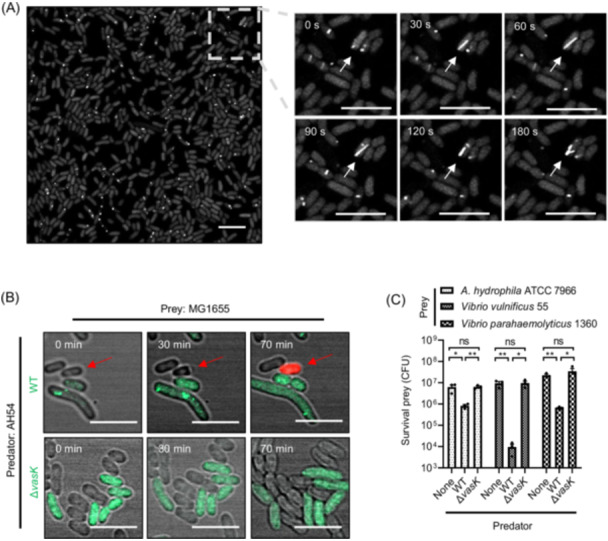
Aeromonas hydrophila, an opportunistic pathogen, often encodes Type VI Secretion System (T6SS) genes. However, the specific functions of T6SS, particularly in the context of clinical strains, remain poorly understood. In this study, we characterize a multi-drug-resistant strain, AH54, which possesses a complete and functional T6SS, composed of a structural cluster and two homologous auxiliary clusters (Aux1 and Aux2). Each auxiliary cluster encodes two distinct effector proteins: a rearrangement hotspot (Rhs) protein and a proline-alanine-arginine repeat (PAAR) protein-Rhs1/PAAR1 in Aux1 and Rhs2/PAAR2 in Aux2. Our findings reveal that AH54 assembles a fully operational T6SS capable of delivering these effectors, driving inter-bacterial antagonism. Interestingly, the T6SS activity in AH54 is temperature-regulated, with enhanced secretion and antibacterial activity at lower temperatures. To protect itself from self-intoxication, AH54 produces immunity proteins (Tsi1-Tsi4) that neutralize the toxic effectors. While PAAR1 and PAAR2 are critical for Hcp secretion, immunity proteins Tsi3 and Tsi4 do not cross-protect against PAAR effectors, suggesting distinct roles for each PAAR protein in optimizing AH54's competitive fitness. In addition, using a Dictyostelium discoideum phagocytosis model, we demonstrate that Rhs2, a metal ion-dependent DNase effector, plays a crucial role in protecting AH54 from eukaryotic predation via T6SS. These findings highlight the pivotal role of T6SS in bacterial competition and pathogenesis, offering new insights into the virulence mechanisms of A. hydrophila.
Keywords: Aeromonas species; T6SS; anti‐eukaryotic virulence; microbial interaction.
© 2025 The Author(s). mLife published by John Wiley & Sons Australia, Ltd on behalf of Institute of Microbiology, Chinese Academy of Sciences.
Conflict of interest statement
The authors declare no conflict of interests.
Figures






References
LinkOut - more resources
Full Text Sources
Miscellaneous