

This is a preprint.
Paclitaxel neurotoxicity is triggered by epidermal EG5-dependent microtubule fasciculation and X-ROS formation
- PMID: 40894047
- PMCID: PMC12393485
- DOI: 10.21203/rs.3.rs-5470731/v1
Paclitaxel neurotoxicity is triggered by epidermal EG5-dependent microtubule fasciculation and X-ROS formation
Abstract
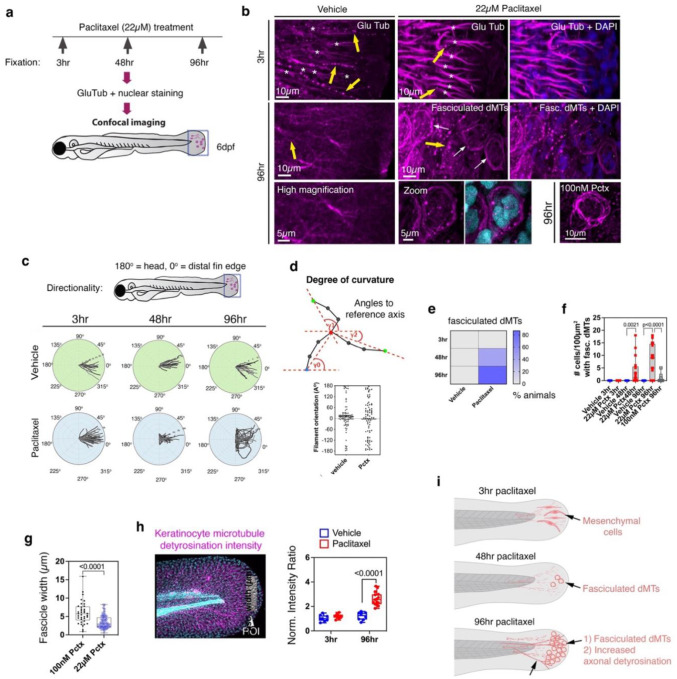
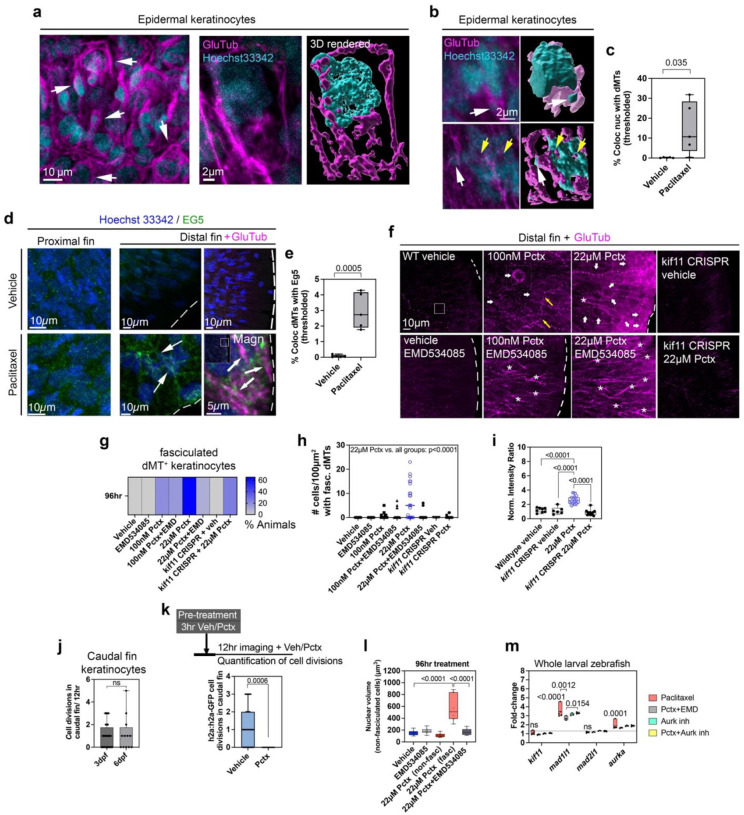
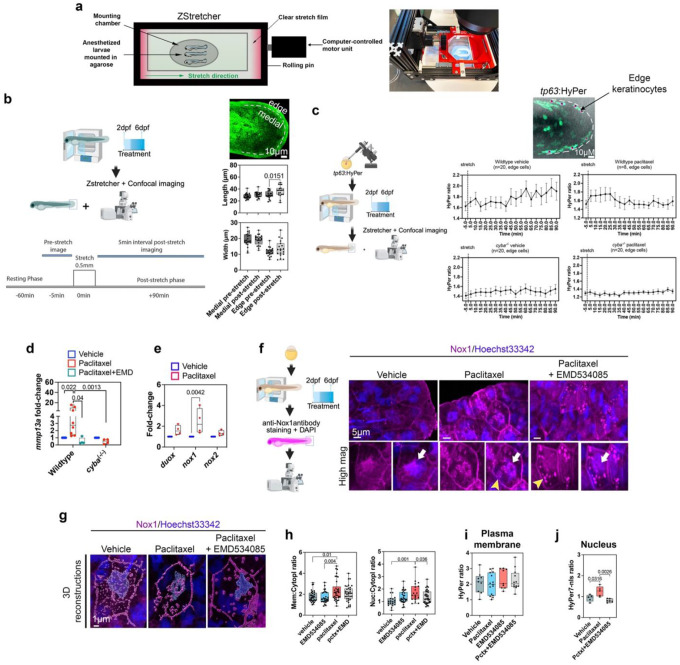
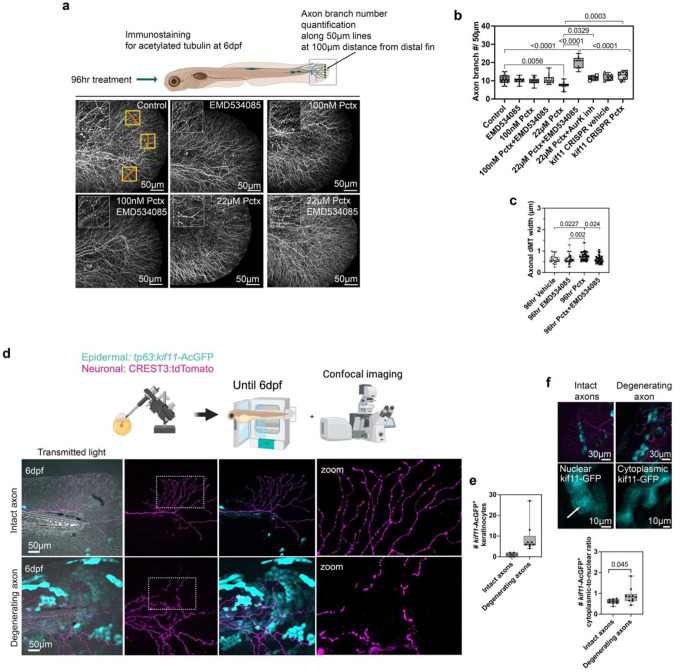
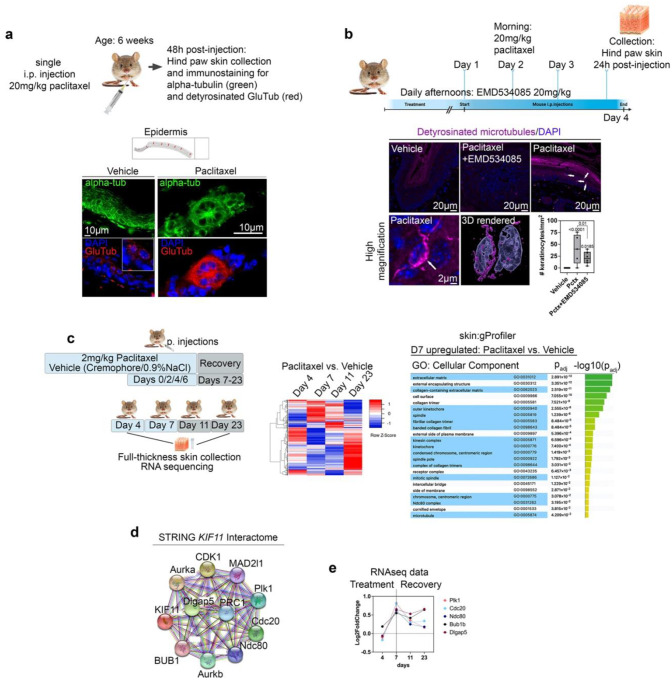
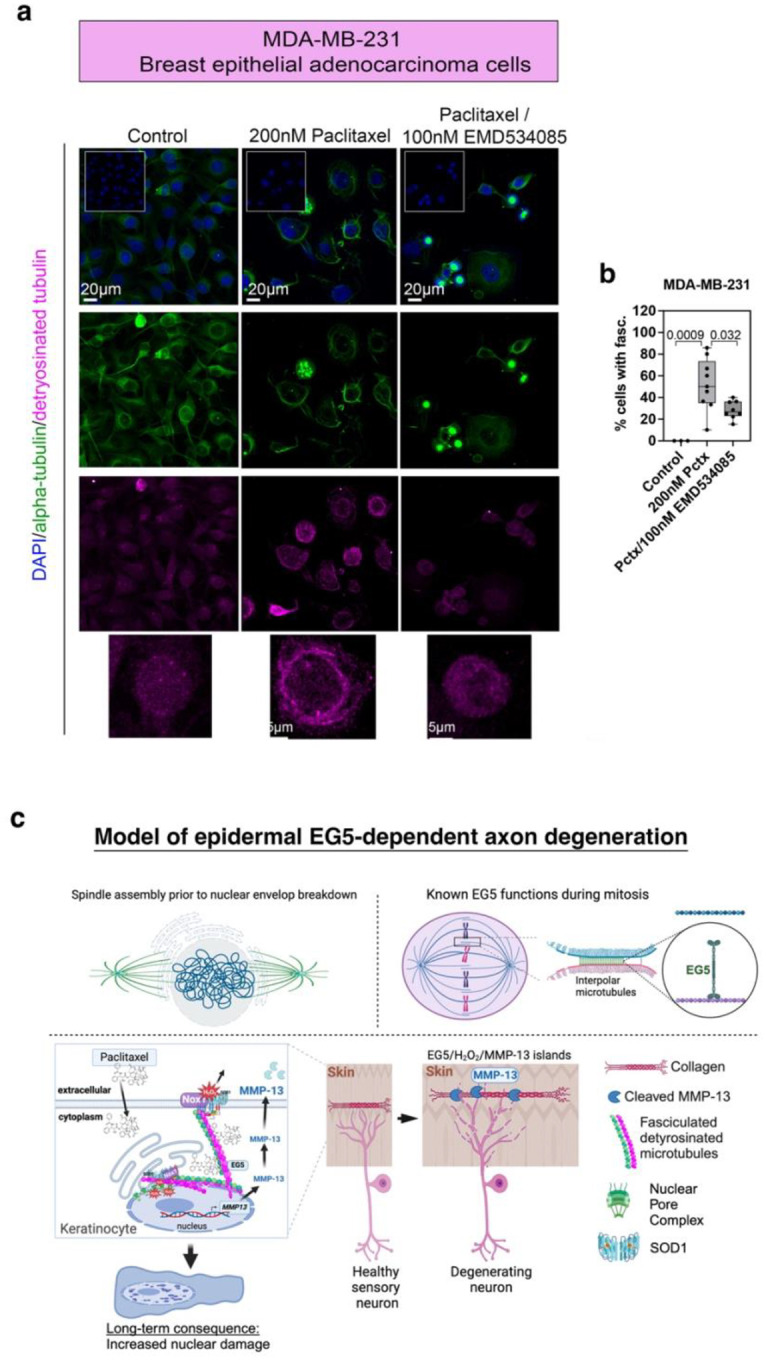
Taxanes are frontline chemotherapeutics that stabilize microtubules, induce mitotic arrest, and drive tumor remission. However, their off-target effects in healthy tissues, most notably cutaneous axon degeneration underlying chemotherapy-induced peripheral neuropathy (CIPN), remain poorly understood. Here, we show that paclitaxel induces microtubule fasciculation in epidermal keratinocytes through the mitotic kinesin Eg5, thereby initiating CIPN. Mechanistically, paclitaxel enhances Eg5-dependent fasciculation of detyrosinated (stabilized) microtubules, which constrict and breach the nuclear lamina. This deformation triggers tension-dependent NADPH oxidase-mediated nuclear ROS (X-ROS) formation upstream of mmp13 transcription, a pathway we previously demonstrated drives sensory axon degeneration. Employing a cross-species framework spanning zebrafish, mice, human skin biopsies, and a breast adenocarcinoma cell line, we uncover a conserved paclitaxel-Eg5 mechanism leading to fasciculation of stable microtubules in both healthy epidermis and cancer cells. These findings highlight the dualistic nature of paclitaxel action and underscore the challenge of preserving anticancer efficacy while preventing neurotoxic side effects.
Keywords: C57BL; Eg5; Kif11; Kinesin-5; MMP13; Taxol; cell cycle; detyrosination; fasciculation; keratinocyte; mice; microtubules; nucleus; paclitaxel; patient; peripheral neuropathy; zebrafish.
Conflict of interest statement
Competing interests. The authors have no competing interests in the research.
Figures
Similar articles
-
Prescription of Controlled Substances: Benefits and Risks.2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 30726003 Free Books & Documents.
-
A rapid and systematic review of the clinical effectiveness and cost-effectiveness of topotecan for ovarian cancer.Health Technol Assess. 2001;5(28):1-110. doi: 10.3310/hta5280. Health Technol Assess. 2001. PMID: 11701100
-
Paclitaxel-induced mitotic arrest results in a convergence of apoptotic dependencies that can be safely exploited by BCL-XL degradation to overcome cancer chemoresistance.bioRxiv [Preprint]. 2025 Jun 26:2025.06.24.661170. doi: 10.1101/2025.06.24.661170. bioRxiv. 2025. PMID: 40666988 Free PMC article. Preprint.
-
Taxane monotherapy regimens for the treatment of recurrent epithelial ovarian cancer.Cochrane Database Syst Rev. 2022 Jul 12;7(7):CD008766. doi: 10.1002/14651858.CD008766.pub3. Cochrane Database Syst Rev. 2022. PMID: 35866378 Free PMC article.
-
Exploring clinical markers of Axon degeneration processes in Chemotherapy-induced peripheral neuropathy among young adults receiving vincristine or paclitaxel.BMC Neurol. 2024 Sep 28;24(1):366. doi: 10.1186/s12883-024-03877-9. BMC Neurol. 2024. PMID: 39342135 Free PMC article.
References
-
- Argyriou A. A., Zolota V., Kyriakopoulou O. & Kalofonos H. P. Toxic peripheral neuropathy associated with commonly used chemotherapeutic agents. J BUON 15, 435–446 (2010). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
