

This is a preprint.
Evolution of Cajal-Retzius Cells in Vertebrates from an Ancient Class of Tp73+ Neurons
- PMID: 40894618
- PMCID: PMC12393536
- DOI: 10.1101/2025.08.21.671508
Evolution of Cajal-Retzius Cells in Vertebrates from an Ancient Class of Tp73+ Neurons
Abstract
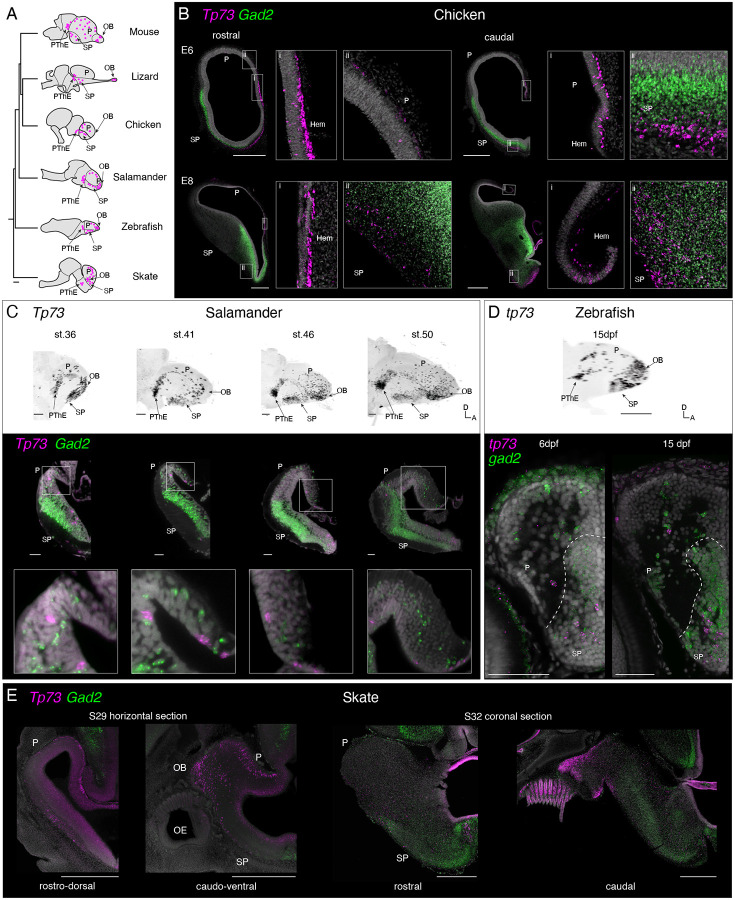
In the developing cerebral cortex, Cajal Retzius (CR) cells are early-born neurons that orchestrate the development of mammalian-specific cortical features. However, this cell type has not been conclusively identified in non-mammalian species. Here we studied neurons expressing Tp73, a transcription factor specifically expressed in most mammalian CR cells. Comparisons of chicken, salamander, zebrafish, and little skate data indicate that Tp73-expressing neurons have conserved spatial distribution and transcriptomic signatures in vertebrates. Among the conserved Tp73-expressing cell types we find CR cells andTp73+ external tufted cells (ETCs) in the olfactory bulb of jawed vertebrates. ETCs and CR cells share the expression of most "canonical" CR cell transcription factors, such as Tp73, Lhx1, Lhx5, Ebf3, and Nr2f2, indicating that they are sister cell types. Our findings suggest that CR and ETCs evolved in stem vertebrates from cells involved in olfactory processing, with CR cells progressively acquiring new specialized roles in developmental signaling.
Figures




References
-
- Goffinet A. M., The evolution of cortical development: the synapsid-diapsid divergence. Development 144, 4061–4077 (2017). - PubMed
-
- Elorriaga V., Pierani A., Causeret F., Cajal-retzius cells: Recent advances in identity and function. Curr. Opin. Neurobiol. 79, 102686 (2023). - PubMed
-
- Kirischuk S., Luhmann H. J., Kilb W., Cajal-Retzius cells: update on structural and functional properties of these mystic neurons that bridged the 20th century. Neuroscience 275, 33–46 (2014). - PubMed
-
- Causeret F., Moreau M. X., Pierani A., Blanquie O., The multiple facets of Cajal-Retzius neurons. Development 148, dev199409 (2021). - PubMed
Publication types
LinkOut - more resources
Full Text Sources