

This is a preprint.
Toggling of NKG2A expression drives functional specialization of iPSC-derived CAR NK cells
- PMID: 40894717
- PMCID: PMC12393553
- DOI: 10.1101/2025.08.20.671199
Toggling of NKG2A expression drives functional specialization of iPSC-derived CAR NK cells
Abstract
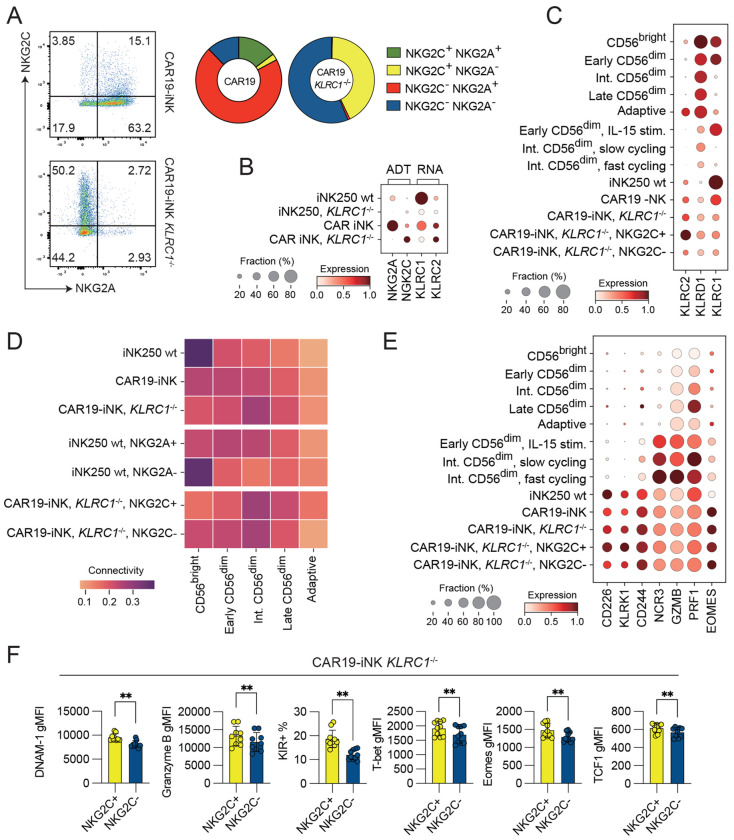
Induced pluripotent stem cell (iPSC)-derived natural killer (iNK) cells offer a promising platform for off-the-shelf immunotherapy against hematological malignancies. NK cell function is dynamically regulated through education driven by inhibitory receptors, including CD94/NKG2A and killer cell immunoglobulin-like receptors (KIR). However, the acquisition of inhibitory receptors in iNK cells and their role during differentiation and education remains poorly defined. In this study, we monitored receptor repertoires, transcriptional states, and functional responses in a range of genetically engineered iNK cell lines. Transcriptional reference mapping placed iNK cells close to cytokine-activated NKG2A+ CD56dim peripheral blood (PB) NK cells. Despite their early differentiation stage, iNK cells displayed a well-developed cytotoxic effector program, which was also reflected in high protein expression of Eomes, granzyme B, and activating receptors DNAM-1 and NKG2D. Acquisition of NKG2A by iNK cells was associated with a more differentiated transcriptional state and superior functional responses against a broad range of targets, including those expressing low to moderate levels of HLA-E, suggesting attenuated inhibitory signaling through NKG2A in iNKs. CRISPR knockout of β2-microglobulin (B2M) in iNK cells revealed that the functional potency of NKG2A+ iNK cells was independent of educating interactions with HLA-E in cis or trans. Finally, CRISPR-mediated ablation of NKG2A led to a spontaneous compensatory surface expression of CD94/NKG2C heterodimers, associated with enhanced IFN-γ production and cytotoxic activity against target cells with forced high expression of single-chain β2m-HLA-E-peptide trimers. Our results indicate an education-independent functional maturation of iNK cells, characterized by potent effector programs coupled with a favorable early-stage transcriptional profile.
Figures





References
-
- Saetersmoen M.L., Hammer Q., Valamehr B., Kaufman D.S. & Malmberg K.J. Off-the-shelf cell therapy with induced pluripotent stem cell-derived natural killer cells. Semin Immunopathol 41, 59–68 (2019). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous