

RNA trans-splicing to rescue β-catenin: A novel approach for treating CTNNB1-Haploinsufficiency disorder
- PMID: 40896583
- PMCID: PMC12398835
- DOI: 10.1016/j.omtn.2025.102680
RNA trans-splicing to rescue β-catenin: A novel approach for treating CTNNB1-Haploinsufficiency disorder
Abstract
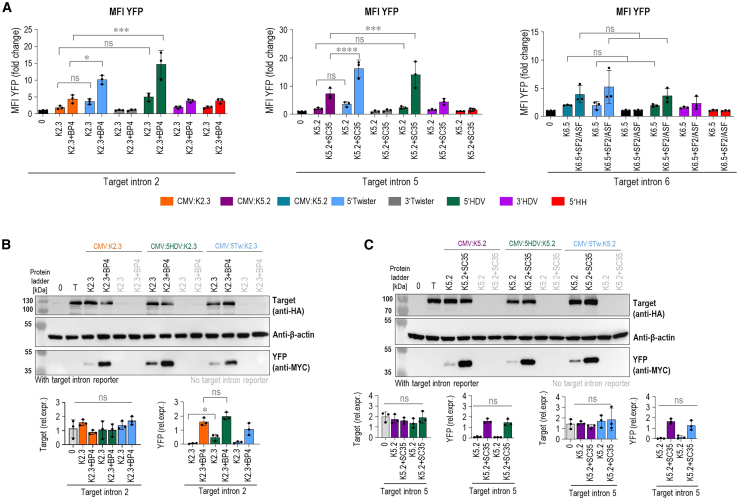
Loss-of-function mutations in the CTNNB1 gene cause β-catenin deficiency, resulting in CTNNB1 syndrome, a rare neurodevelopmental disorder characterized by motor and cognitive impairments. Given the wide variety of mutations across CTNNB1 and its dosage sensitivity, a mutation-independent therapeutic approach that preserves endogenous gene regulation is critically needed. This study introduces spliceosome-mediated RNA trans-splicing as a novel approach to restore β-catenin production. Pre-trans-splicing RNA molecules (PTMs) targeting CTNNB1 introns 2, 5, and 6 were designed and evaluated using a split yellow fluorescent protein reporter system. Rationally designed short antisense RNAs, which mask splicing regulatory elements, significantly enhanced PTM-mediated trans-splicing at both mRNA and protein levels. Additionally, introducing a self-cleaving ribozyme at the PTM's 5' end further improved trans-splicing efficiency, likely due to increased nuclear retention. CMV promoter-driven PTM expression yielded the highest efficiency. Importantly, successful trans-splicing of the endogenous CTNNB1 transcript confirmed the physiological relevance of this strategy. This study is the first to apply and optimize spliceosome-mediated RNA trans-splicing (SMaRT) for CTNNB1 mRNA correction, providing a promising, mutation-agnostic approach for treating CTNNB1 syndrome.
Keywords: CTNNB1 syndrome; MT: RNA/DNA Editing; RNA therapy; rare disease; ribozymes; small antisense RNA; trans-splicing.
© 2025 The Authors.
Conflict of interest statement
The authors declare no competing interests.
Figures







References
-
- López-Rivera J.A., Pérez-Palma E., Symonds J., Lindy A.S., McKnight D.A., Leu C., Zuberi S., Brunklaus A., Møller R.S., Lal D. A catalogue of new incidence estimates of monogenic neurodevelopmental disorders caused by de novo variants. Brain J. Neurol. 2020;143:1099–1105. doi: 10.1093/brain/awaa051. - DOI - PMC - PubMed
-
- de Ligt J., Willemsen M.H., van Bon B.W.M., Kleefstra T., Yntema H.G., Kroes T., Vulto-van Silfhout A.T., Koolen D.A., de Vries P., Gilissen C., et al. Diagnostic Exome Sequencing in Persons with Severe Intellectual Disability. N. Engl. J. Med. 2012;367:1921–1929. doi: 10.1056/NEJMoa1206524. - DOI - PubMed
-
- Verhoeven W.M.A., Egger J.I.M., Jongbloed R.E., van Putten M.M., de Bruin-van Zandwijk M., Zwemer A.-S., Pfundt R., Willemsen M.H. A de novo CTNNB1 Novel Splice Variant in an Adult Female with Severe Intellectual Disability. Int. Med. Case Rep. J. 2020;13:487–492. doi: 10.2147/IMCRJ.S270487. - DOI - PMC - PubMed
-
- Kuechler A., Willemsen M.H., Albrecht B., Bacino C.A., Bartholomew D.W., van Bokhoven H., van den Boogaard M.J.H., Bramswig N., Büttner C., Cremer K., et al. De novo mutations in beta-catenin (CTNNB1) appear to be a frequent cause of intellectual disability: expanding the mutational and clinical spectrum. Hum. Genet. 2015;134:97–109. doi: 10.1007/s00439-014-1498-1. - DOI - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
