

ATE1 promotes breast cancer progression via arginylation-dependent regulation of MAPK-MYC signaling
- PMID: 40898325
- PMCID: PMC12403503
- DOI: 10.1186/s12964-025-02376-9
ATE1 promotes breast cancer progression via arginylation-dependent regulation of MAPK-MYC signaling
Abstract
Background: Arginyl-tRNA-protein transferase (ATE1) catalyzes N-terminal arginylation, a regulatory protein modification implicated in various cellular processes, including proliferation, apoptosis, and migration. Although ATE1 has context-dependent roles in cancer, its specific function in breast cancer remains unclear. This study investigates the oncogenic role of ATE1 across multiple breast cancer subtypes and its underlying molecular mechanisms.
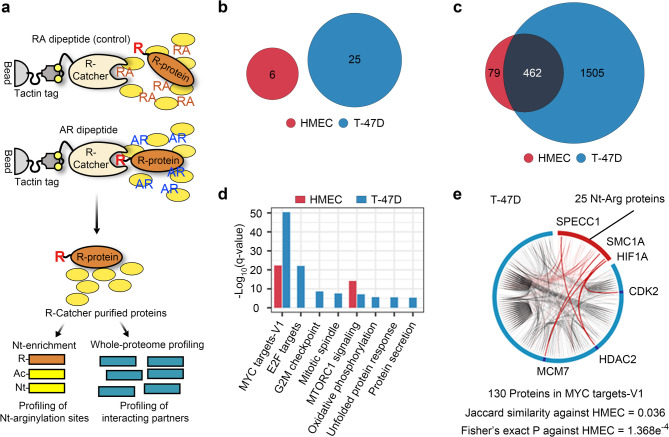
Methods: ATE1 expression in breast cancer was evaluated using TCGA data and immunoblotting across breast cancer cell lines and normal mammary epithelial cells (HMEC). Functional studies using siRNA- and shRNA-mediated knockdown assessed ATE1's role in cell viability, clonogenic growth, migration, and tumorigenesis in vitro and xenograft models. Quantitative proteomics, R-catcher-based N-terminomics, and pathway analyses were employed to identify ATE1-dependent signaling networks, with a focus on MAPK-MYC axis regulation. Flow cytometry and immunoblotting were used to assess cell cycle progression, apoptosis, and MYC stability.
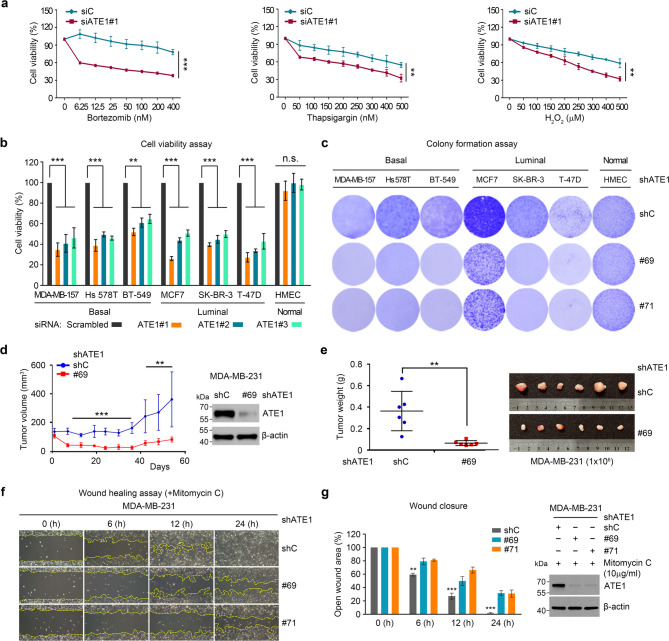
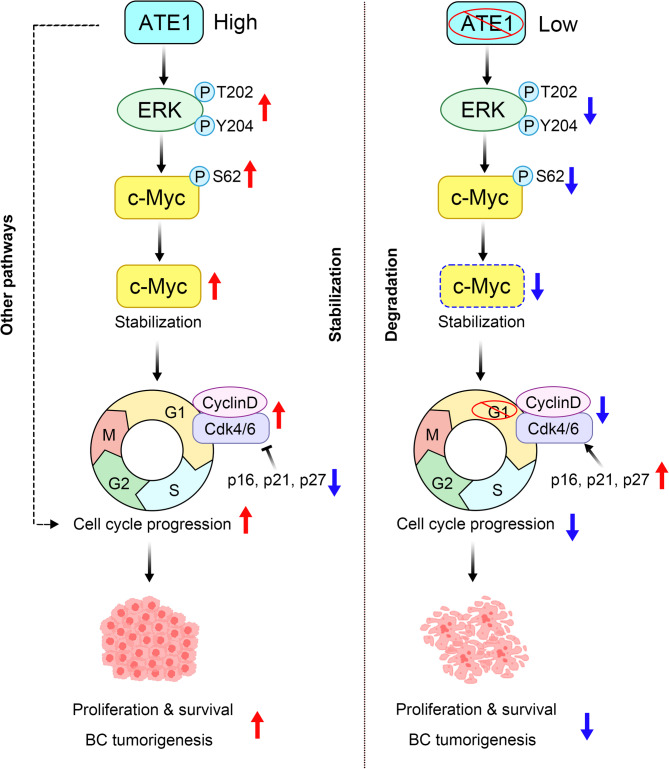
Results: ATE1 was significantly upregulated in breast cancer cells and associated with poor prognosis in early-stage patients. ATE1 depletion selectively impaired viability, proliferation, and migration in breast cancer cells, but not in HMECs. In vivo, ATE1 silencing suppressed tumor growth in xenograft models. Proteomic profiling revealed that ATE1 regulates the cell cycle and survival pathways in a subtype-specific manner, particularly through modulation of the MAPK-MYC-CDK6 axis in luminal T-47D cells. ATE1 stabilized MYC protein via ERK-mediated phosphorylation at Ser62, promoting cell cycle progression and suppressing apoptosis. Rescue experiments confirmed that ATE1's tumor-promoting activity depends on its arginyltransferase function.
Conclusions: ATE1 promotes breast cancer progression by enhancing cell proliferation, survival, and migration through MAPK-dependent stabilization of MYC in a lineage-specific context. These findings identify ATE1 as a potential therapeutic target and highlight the relevance of protein arginylation in the molecular heterogeneity of breast cancer.
Keywords: ATE1 (arginyltransferase 1); Breast cancer; Cell proliferation; Cell survival; MYC signaling; N-degron pathway; N-terminal arginylation; Tumor progression.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: All animal experiments were approved by the Animal Care and Ethics Committee of the Korea Research Institute of Bioscience and Biotechnology (Approval No. KRIBB-AEC-14124; Daejeon, Korea), which also provided oversight and support for this study. Consent for publication: Not applicable. Competing interests: The authors declare no competing interests.
Figures




References
MeSH terms
Substances
Grants and funding
- KGM1062413/The Korea Research Institute of Bioscience and Biotechnology (KRIBB) Research Initiative Program
- RS-2023-00279134/Ministry of Science and ICT
- GTL24021-000/National Research Council of Science & Technology (NST) grant
- RS-2024-00444177/The Nano & Material Technology Development Program through the National Research Foundation of Korea (NRF)
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
