

Insights Into the Almond Domestication History
- PMID: 40901495
- PMCID: PMC12399410
- DOI: 10.1111/eva.70150
Insights Into the Almond Domestication History
Abstract
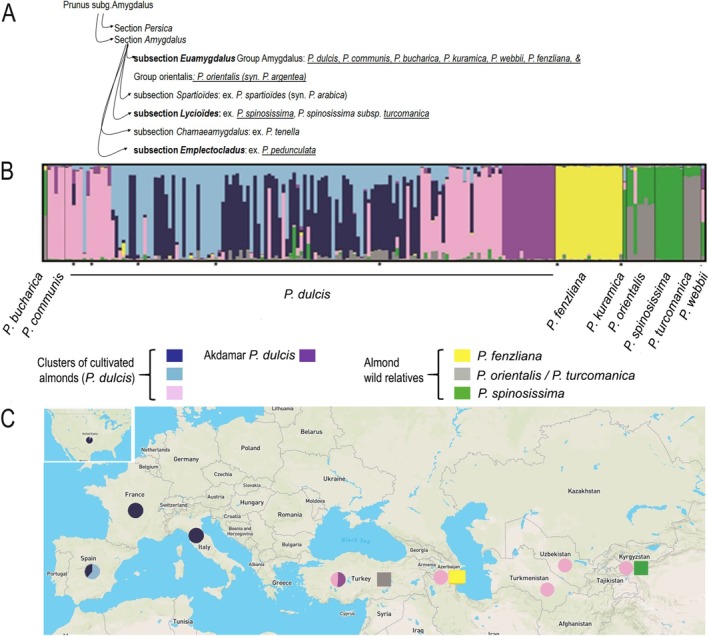
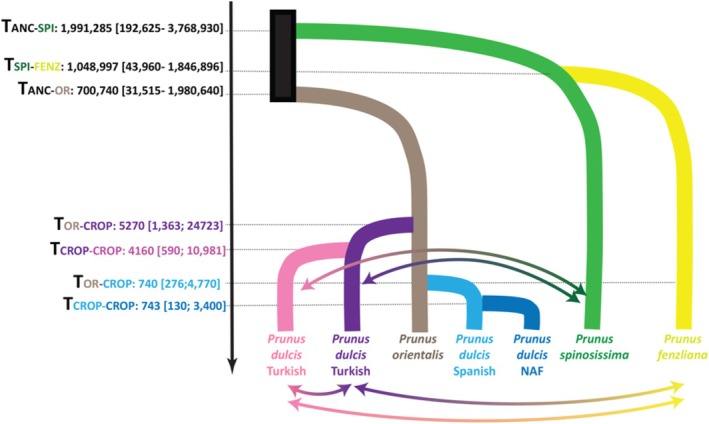
Understanding crop domestication offers crucial insights into the evolutionary processes that drive population divergence and adaptation. It also informs the identification of genetically diverse wild germplasm, which is essential for breeding and conservation efforts. While domestication has been extensively studied in many Mediterranean fruit trees, the evolutionary history of the almond (Prunus dulcis) remains comparatively underexplored. To address this, we analyzed 209 wild and cultivated almond accessions sampled across Eurasia and genotyped with 23 microsatellite markers. Using population genetics and coalescent-based inference, we reconstructed the domestication history of P. dulcis and its relationships with wild relatives. Bayesian clustering revealed four genetically distinct clusters of cultivated almonds: Turkish, Caucasian-Central Asian, Southern Spanish, and European/North American. These groups were differentiated from wild almond species-including Prunus turcomanica, Prunus orientalis, Prunus fenzliana, and Prunus spinosissima-each forming its gene pool across the Middle East and Central Asia. Approximate Bayesian Computation (ABC) supported a single domestication event in the Middle East, originating from either P. orientalis or P. turcomanica, with subsequent gene flow from P. fenzliana and P. spinosissima into the Turkish and Central Asian cultivated gene pools, respectively. We also inferred reciprocal introgression from cultivated almonds back into wild populations. Notably, sharka resistance-caused by plum pox virus (PPV)-was identified in three P. dulcis clusters and P. fenzliana, suggesting that resistance may have arisen independently or been maintained through crop-wild introgression. Together, our results highlight a complex and protracted domestication history for almond, shaped by contributions from multiple wild relatives and recurrent gene flow. These findings enhance our understanding of perennial crop evolution and underscore the value of wild germplasm in breeding programs aimed at increasing resilience in fruit trees.
Keywords: Mediterranean Basin; Prunus; almonds; domestication; fruit trees; gene flow; genetic resources; sharka; virus.
© 2025 The Author(s). Evolutionary Applications published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures


References
-
- Abbo, S. , Gopher A., and Lev‐Yadun S.. 2015. “Fruit Domestication in the Near East.” Plant Breeding Reviews 39: 325–378.
-
- Ak, B. E. , Açar I., and Sakar E.. 2001. “An Investigation on the Determination of Pomological and Morphological Traits of Wild Almond Grown at Sanliurfa Province.” In XI GREMPA Seminar on Pistachios and Almonds, edited by Ak B. E., vol. 56, 139–144. CIHEAM.
-
- Belkhir, K. , Borsa P., Chikhi L., Raufaste N., and Bonhomme F.. 2004. “GENETIX4. 05, Logiciel sous Windows TM pour la génétique des Populations.” Laboratoire Génome, Populations, Interactions, CNRS UMR 5000 Université de Montpellier II, 5000.
LinkOut - more resources
Full Text Sources
