

The ILR3-NRTs/NIA1/SWEET12 module regulates nitrogen uptake and utilization in apple
- PMID: 40903771
- PMCID: PMC12406481
- DOI: 10.1186/s43897-025-00172-0
The ILR3-NRTs/NIA1/SWEET12 module regulates nitrogen uptake and utilization in apple
Abstract
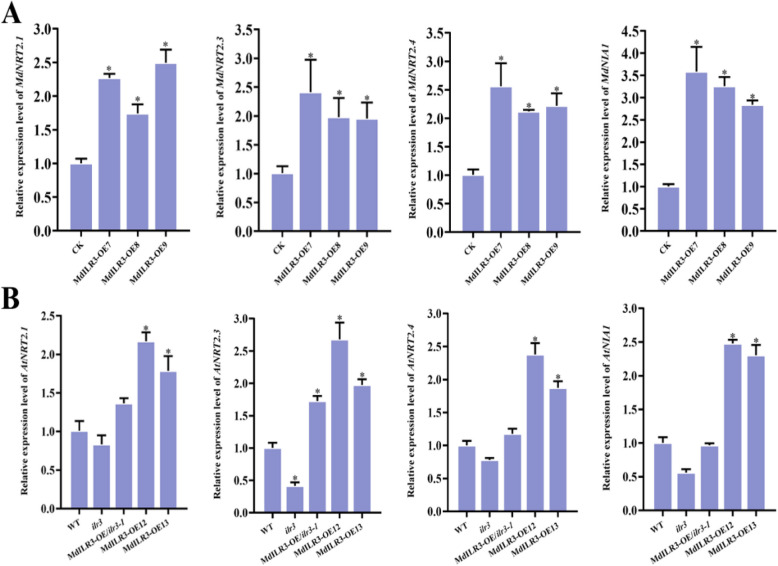
Nitrogen (N) is essential for the physiological metabolism, growth, and development of plants. Plants have evolved a complex regulatory network for the efficient regulation of N uptake and utilization to adapt to fluctuations in environmental N levels. However, the mechanisms underlying the regulation of N absorption and utilization in apple remain unclear. Here, we identified MdILR3 (IAA-LEUCINE RESISTANT3) as an upstream regulator of MdNRT2.4 through yeast one-hybrid (Y1H) screening. MdILR3 overexpression significantly up-regulated the expression of MdNRT2.3/2.4 and MdNIA1, resulting in an increase in nitrate content and nitrate reductase activity. Y1H and EMSA assays revealed that MdILR3 directly interacted with the promoters of MdNRT2.3/2.4 and MdNIA1. Furthermore, MdILR3 can directly bind to the promoter of MdSWEET12 and activate its expression, thereby regulating sucrose transport to provide energy for N uptake in roots. In summary, we provide physiological and molecular evidence suggesting that MdILR3 may positively regulate nitrate response by activating the expression of genes related to N uptake and sugar transport. Our findings suggest that genetic improvements in apple could enhance its ability to absorb and utilize N.
Keywords: MdILR3 (IAA-LEUCINE RESISTANT3); Nitrate assimilation; Nitrate transporter; Nitrogen use efficiency; Sugar transport.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: Not applicable. Consent for publication: All authors approve the manuscript and consent to the publication of the work. Competing interests: The authors declare that there are no competing interests.
Figures







References
-
- Ayre BG. Membrane-transport systems for sucrose in relation to whole-plant carbon partitioning. Mol Plant. 2011;4:377–94. - PubMed
-
- Bellegarde F, Gojon A, Martin A. Signals and players in the transcriptional regulation of root responses by local and systemic N signaling in Arabidopsis thaliana. J Exp Bot. 2017;68:2553–65. - PubMed
-
- Bouguyon E, Brun F, Meynard D, Kubes M, Pervent M, Leran S, et al. Multiple mechanisms of nitrate sensing by Arabidopsis nitrate transceptor NRT1.1. Nat Plants. 2015;1:15015. - PubMed
Grants and funding
- 32372642/National Natural Science Foundation of China
- 32272683/National Natural Science Foundation of China
- 2022YFD1201700/National Key Research and Development Program of China
- ZR2024JQ036/National Natural Science Foundation of Shandong Province
- CARS-27/China Agriculture Research System of MOF and MARA
LinkOut - more resources
Full Text Sources