

Gut microbiota dysbiosis affects intestinal sensitivity through epithelium-to-neuron signaling: novel insights from a colon organoid-based model to improve visceral pain therapy
- PMID: 40903878
- PMCID: PMC12413070
- DOI: 10.1080/19490976.2025.2547029
Gut microbiota dysbiosis affects intestinal sensitivity through epithelium-to-neuron signaling: novel insights from a colon organoid-based model to improve visceral pain therapy
Abstract
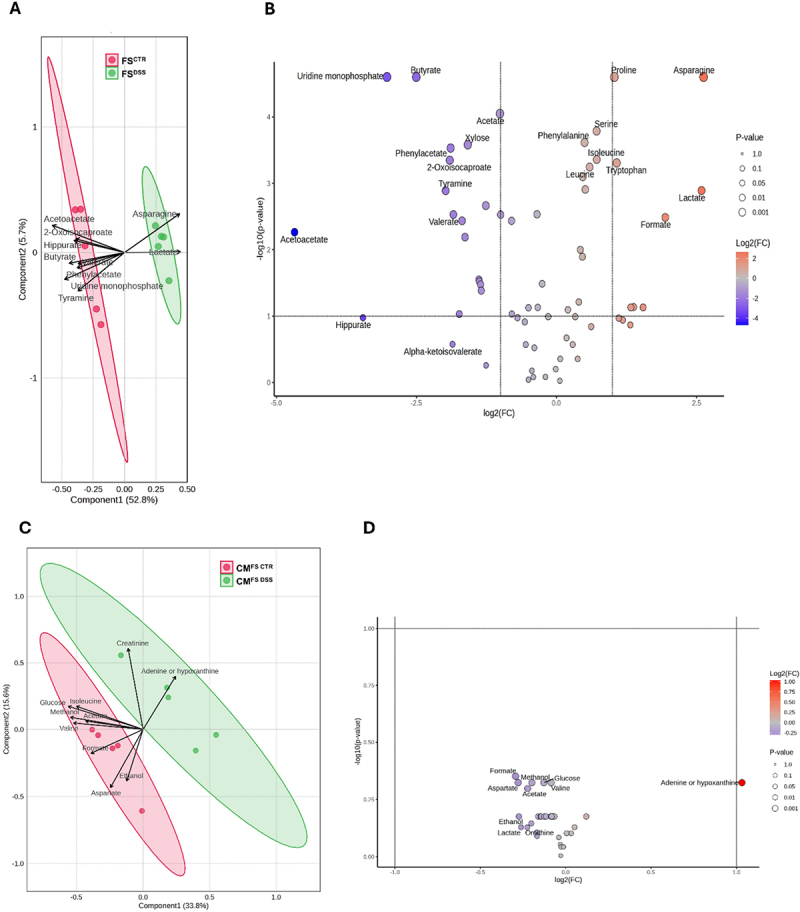
Chronic gastrointestinal pain is a hallmark of most intestinal pathologies, yet effective treatments remain elusive given the complexity of the underlying mechanisms. Aiming to investigate the intestinal epithelium contribution to visceral pain modulation in dysbiosis context, we first demonstrated that intracolonic instillation of microbe-free fecal supernatants from mice with post-inflammatory dysbiosis induced by dextran sodium sulfate (FSDSS) provokes visceral hypersensitivity in recipient mice. Epithelium involvement in the response to FSDSS was analyzed through a novel in vitro approach comprising murine epithelial colon organoids and primary dorsal root ganglia (DRG) neurons. FSDSS treatment induced growth and metabolic impairment in colon organoids, which revealed a dysbiosis-driven epithelial dysfunction. Notably, the combination of FSDSS and conditioned medium from FSDSS-treated colon organoids induced an increase in DRG neuron intrinsic excitability, along with greater immunoreactivity to c-Fos and calcitonin-gene related peptide, implicating an integrated role of both microbial and epithelial products in visceral sensitivity regulation. By investigating the underlying signaling, metabolomic analysis revealed reduced levels of short chain fatty acids in FSDSS, such as butyrate, acetate, valerate, and propionate. Moreover, transcriptomic analysis of FSDSS-treated colon organoids showed the dysregulated expression of several signaling factors by which intestinal epithelium may modulate sensory neuron excitability, including proteases, cytokines, neuromodulators, growth factors, and hormones. These findings provide novel insights into the role of gut epithelium in the modulation of sensory neuron excitability under dysbiosis conditions, emphasizing that targeting epithelial-neuronal signaling might represent a promising therapeutic strategy for visceral pain management.
Keywords: DRG neurons; Visceral pain; dysbiosis; epithelial-neuronal signaling; intestinal epithelium; microbiota; organoids.
Conflict of interest statement
No potential conflict of interest was reported by the author(s).
Figures







References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources