

This is a preprint.
Sequencing of distinct wing behaviors during Drosophila courtship
- PMID: 40909675
- PMCID: PMC12407689
- DOI: 10.1101/2025.08.21.671456
Sequencing of distinct wing behaviors during Drosophila courtship
Update in
-
Sequencing of distinct wing behaviors during Drosophila courtship.Curr Biol. 2026 Feb 18:S0960-9822(26)00078-3. doi: 10.1016/j.cub.2026.01.042. Online ahead of print. Curr Biol. 2026. PMID: 41713408
Abstract
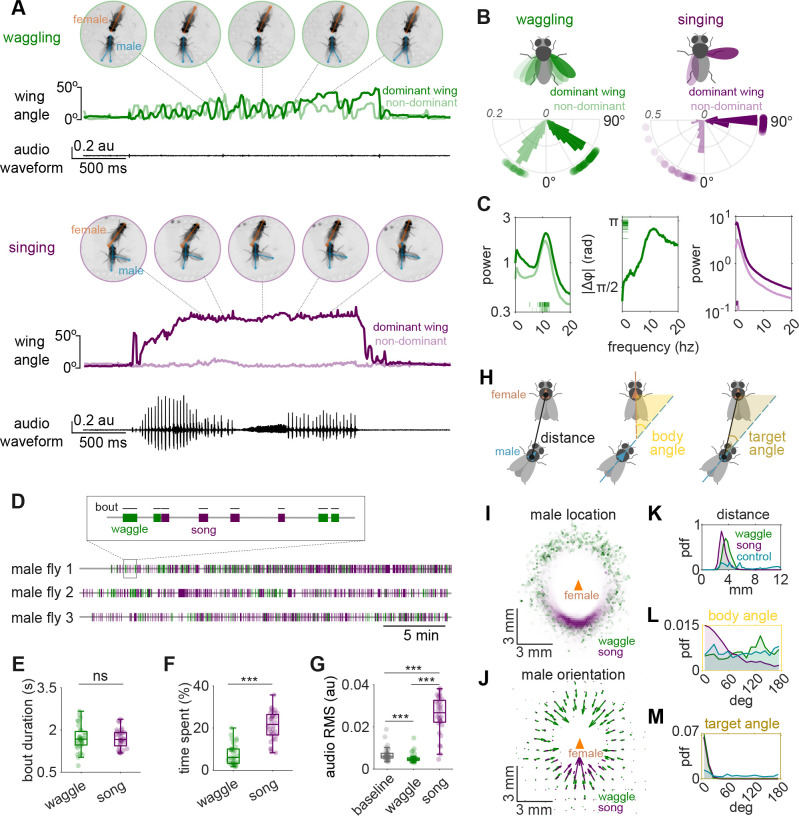
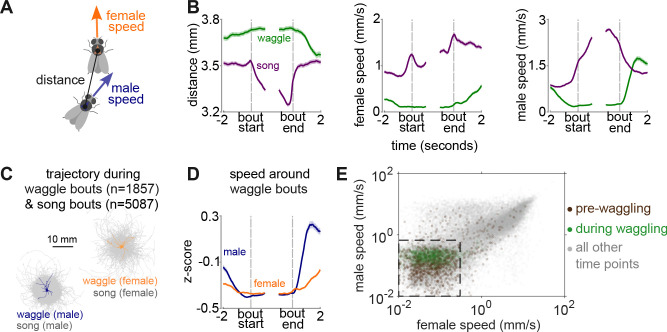
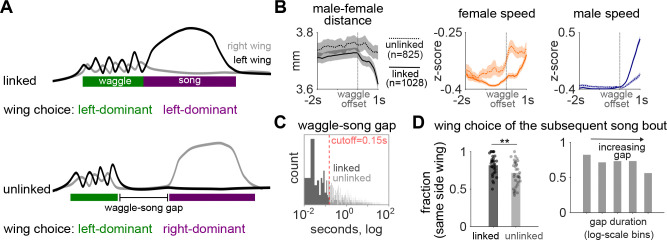
Some behaviors, like biting followed by chewing and then swallowing, unfold in stereotyped sequences, while others, such as limb movements during defensive maneuvers, can be flexibly combined as needed. During courtship, male Drosophilid flies produce a series of actions, including orientation, tapping, singing, licking, and copulation, that follow an ordered but temporally variable sequence [1,2]. At shorter timescales, however, individual actions remain highly dynamic. For example, courtship songs are composed of variable sequences of distinct syllables, with their patterning and amplitude actively shaped by female cues [3-5]. Leveraging recent advances in behavioral quantification [6], we discover a new courtship wing behavior that we term "waggling", which is present across multiple Drosophila species and characterized by rhythmic, anti-phase wing movements. We identify an intermediate level of stereotyped behavioral structure: a directional three-part motif where males and females first decelerate to near-complete stillness, followed by male-initiated waggling, which then transitions into courtship song. Wing kinematics during waggle bouts are predictive of wing choice in subsequent songs, suggesting waggling may serve as a preparatory behavior. We then focus on P1/pC1 neurons, known to promote courtship [5,7-11]. Optogenetic activation of specific P1/pC1 neuron subsets in solitary males, without any female cues, is sufficient to recapitulate the entire stillness-to-waggling-to-singing progression. These findings reveal a new layer of stereotyped structure within a flexible courtship display and demonstrate that P1/pC1 neurons can orchestrate multi-action behavioral programs through internal dynamics.
Keywords: Drosophila; behavioral sequencing; courtship; locomotion; neuroethology; social interactions.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures


References
-
- Anderson DJ, Perona P. Toward a science of computational ethology. Neuron. 2014;84: 18–31. - PubMed
-
- Yamamoto D, Koganezawa M. Genes and circuits of courtship behaviour in Drosophila males. Nat Rev Neurosci. 2013;14: 681–692. - PubMed
-
- Coen P, Clemens J, Weinstein AJ, Pacheco DA, Deng Y, Murthy M. Dynamic sensory cues shape song structure in Drosophila. Nature. 2014;507: 233–237. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous