

Platelet factor 4 regulates hematopoietic stem cell aging
- PMID: 40920871
- PMCID: PMC12574973
- DOI: 10.1182/blood.2024027432
Platelet factor 4 regulates hematopoietic stem cell aging
Abstract
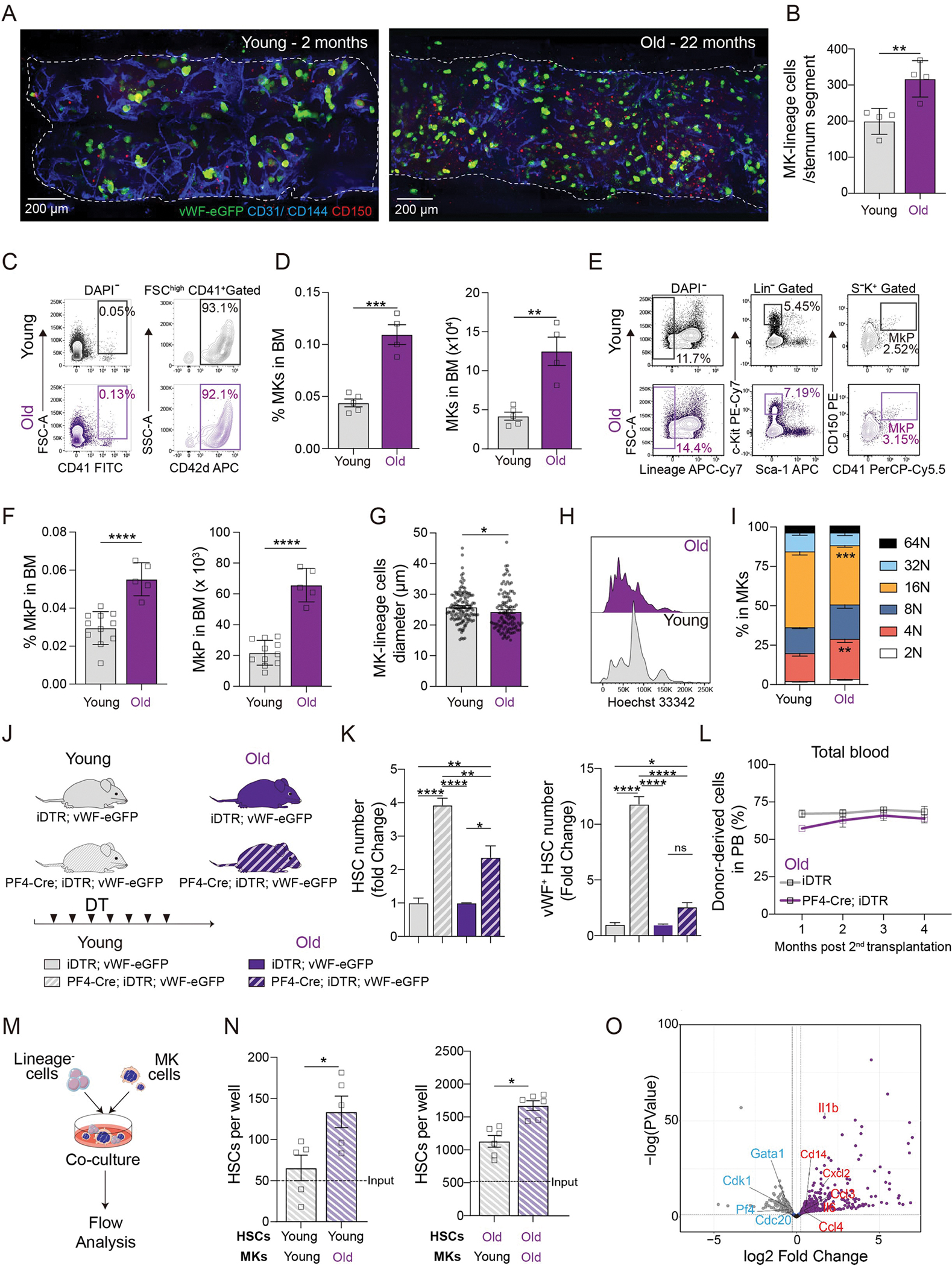
Hematopoietic stem cells (HSCs) responsible for blood cell production and their bone marrow regulatory niches undergo age-related changes, affecting immune responses and predisposing individuals to hematologic malignancies. Here, we show that the age-related alterations of the megakaryocytic niche and associated downregulation of platelet factor 4 (PF4) are pivotal mechanisms driving HSC aging. PF4-deficient mice display several phenotypes reminiscent of accelerated HSC aging, including lymphopenia, increased myeloid output, and DNA damage, mimicking physiologically aged HSCs. Remarkably, recombinant PF4 administration restored old HSCs to youthful functional phenotypes characterized by improved cell polarity, reduced DNA damage, enhanced in vivo reconstitution capacity, and balanced lineage output. Mechanistically, we identified low-density lipoprotein receptor and C-X-C motif chemokine receptor 3 as HSC receptors transmitting the PF4 signal, with double knockout mice exhibiting exacerbated HSC aging phenotypes similar to PF4-deficient mice. Furthermore, human HSCs across various age groups also respond to the youthful PF4 signaling, highlighting its potential for rejuvenating aged hematopoietic systems. These findings pave the way for targeted therapies aimed at reversing age-related HSC decline, with potential implications in the prevention or improvement of the course of age-related hematopoietic diseases.
© 2025 American Society of Hematology. Published by Elsevier Inc. All rights are reserved, including those for text and data mining, AI training, and similar technologies.
Conflict of interest statement
Disclosure of Conflicts of Interest
The authors declare no competing interests.
Figures




Update of
-
Platelet Factor 4 (PF4) Regulates Hematopoietic Stem Cell Aging.bioRxiv [Preprint]. 2024 Nov 27:2024.11.25.625252. doi: 10.1101/2024.11.25.625252. bioRxiv. 2024. Update in: Blood. 2025 Dec 4;146(23):2765-2778. doi: 10.1182/blood.2024027432. PMID: 39651177 Free PMC article. Updated. Preprint.
Comment in
-
The secret to staying young: platelet factor 4.Blood. 2025 Dec 4;146(23):2734-2736. doi: 10.1182/blood.2025031130. Blood. 2025. PMID: 41343166 No abstract available.
References
-
- Lopez-Otin C, Blasco MA, Partridge L, Serrano M, Kroemer G. Hallmarks of aging: An expanding universe. Cell. 2023;186(2):243–278. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
